Citation: James T. Cronin, Jerome Goddard II, Amila Muthunayake, Ratnasingham Shivaji. Modeling the effects of trait-mediated dispersal on coexistence of mutualists[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7838-7861. doi: 10.3934/mbe.2020399

| [1] |
I. S. Pearse, E. LoPresti, R. N. Schaeffer, W. C. Wetzel, K. A. Mooney, J. G. Ali, et al., Generalising indirect defence and resistance of plants, Ecol. Lett., 23 (2020), 1137-1152. doi: 10.1111/ele.13512

|
| [2] |
F. Encinas-Viso, T. A. Revilla, R. S. Etienne, Phenology drives mutualistic network structure and diversity, Ecol. Lett., 15 (2012), 198-208. doi: 10.1111/j.1461-0248.2011.01726.x
|
| [3] |
J. L. Bronstein, Conditional outcomes in mutualistic interactions, Trends Ecol. Evol., 9 (1994), 214-217. doi: 10.1016/0169-5347(94)90246-1
|
| [4] |
J. L. Bronstein, Our current understanding of mutualism, Q Rev. Biol., 69 (1994), 31-51. doi: 10.1086/418432
|
| [5] |
S. Silknetter, R. P. Creed, B. L. Brown, E. A. Frimpong, J. Skelton, B. K. Peoples, Positive biotic interactions in freshwaters: A review and research directive, Freshw. Biol., 65 (2020), 811-832. doi: 10.1111/fwb.13476
|
| [6] |
E. E. Werner, S. D. Peacor, A review of trait-mediated indirect interactions in ecological communities, Ecology, 84 (2003), 1083-1100. doi: 10.1890/0012-9658(2003)084[1083:AROTII]2.0.CO;2
|
| [7] | T. Ohgushi, O. Schmitz, R. D. Holt, Trait-mediated indirect interactions: ecological and evolutionary perspectives, Cambridge University Press, New York, 2012. |
| [8] | R. E. Irwin, The role of trait-mediated indirect interactions for multispecies plant-animal mutualisms, book section 257,257-277, Cambridge University Press, Cambridge, New York., 2012. |
| [9] | R. D. Holt and M. Barfield, Trait-mediated effects, density dependence and the dynamic stability of ecological systems, Cambridge University Press, Cambridge, New York, 2012. |
| [10] |
R. S. Cantrell, C. Cosner, Effects of harvesting mediated by dispersal traits, Nat. Resour. Model., 31 (2018), e12168. doi: 10.1111/nrm.12168
|
| [11] |
J. T. Cronin, K. J. Haynes, F. Dillemuth, Spider effects on planthopper mortality, dispersal, and spatial population dynamics, Ecology, 85 (2004), 2134-2143. doi: 10.1890/03-0591
|
| [12] |
A. Sih, L. B. Kats, R. D. Moore, Effects of predatory sunfish on the density, drift, and refuge use of stream salamander larvae, Ecology, 73 (1992), 1418-1430. doi: 10.2307/1940687
|
| [13] |
H. Hakkarainen, P. Ilmonen, V. Koivunen, E. Korpimäki, Experimental increase of predation risk induces breeding dispersal of tengmalm's owl, Oecologia, 126 (2001), 355-359. doi: 10.1007/s004420000525
|
| [14] |
B. L. Peckarsky, Alternative predator avoidance syndromes of stream-dwelling mayfly larvae, Ecology, 77 (1996), 1888-1905. doi: 10.2307/2265793
|
| [15] |
J. D. Hoeksema, E. M. Bruna, Pursuing the big questions about interspecific mutualism: a review of theoretical approaches, Oecologia, 125 (2000), 321-330. doi: 10.1007/s004420000496
|
| [16] |
W. Völkl, J. Woodring, M. Fischer, M. W. Lorenz, K. H. Hoffmann, Ant-aphid mutualisms: the impact of honeydew production and honeydew sugar composition on ant preferences, Oecologia, 118 (1999), 483-491. doi: 10.1007/s004420050751
|
| [17] |
J. E. Kleinjan and T. E. Mittler, A chemical influence of ants on wing development in aphids, Entomol. Exp. Appl., 18 (1975), 384-388. doi: 10.1111/j.1570-7458.1975.tb00411.x
|
| [18] |
P. Kindlmann, M. Hullé, B. Stadler, Timing of dispersal: effect of ants on aphids, Oecologia, 152 (2007), 625-631. doi: 10.1007/s00442-007-0684-4
|
| [19] |
T. H. Oliver, A. Mashanova, S. R. Leather, J. M. Cook, V. A. A. Jansen, Ant semiochemicals limit apterous aphid dispersal, Proc. Royal Soc. B, 274 (2007), 3127-3131. doi: 10.1098/rspb.2007.1251
|
| [20] |
N. E. Fatouros, G. Bukovinszkine'Kiss, L. A. Kalkers, R. S. Gamborena, M. Dicke, M. Hilker, Oviposition-induced plant cues: do they arrest trichogramma wasps during host location?, Entomol. Exp. Appl., 115 (2005), 207-215. doi: 10.1111/j.1570-7458.2005.00245.x
|
| [21] |
G. E. Heilman, J. R. Strittholt, N. C. Slosser, D. A. Dellasala, Forest fragmentation of the conterminous united states: Assessing forest intactness through road density and spatial characteristics: Forest fragmentation can be measured and monitored in a powerful new way by combining remote sensing, geographic information systems, and analytical software, BioSci., 52 (2002), 411-422. doi: 10.1641/0006-3568(2002)052[0411:FFOTCU]2.0.CO;2
|
| [22] |
R. M. Ewers, R. K. Didham, W. D. Pearse, V. Lefebvre, I. M. D. Rosa, J. M. B. Carreiras, et al., Using landscape history to predict biodiversity patterns in fragmented landscapes, Ecol. Lett., 16 (2013), 1221-1233. doi: 10.1111/ele.12160
|
| [23] |
K. Uchida, A. Ushimaru, Biodiversity declines due to abandonment and intensification of agricultural lands: patterns and mechanisms, Ecol. Monogr., 84 (2014), 637-658. doi: 10.1890/13-2170.1
|
| [24] |
L. Ries, J. Robert, J. Fletcher, J. Battin, T. D. Sisk, Ecological responses to habitat edges: Mechanisms, models, and variability explained, Annu. Rev. Ecol. Evol. Syst., 35 (2004), 491-522. doi: 10.1146/annurev.ecolsys.35.112202.130148
|
| [25] |
W. F. Fagan, R. S. Cantrell, C. Cosner, How habitat edges change species interactions, Am. Nat., 153 (1999), 165-182. doi: 10.1086/303162
|
| [26] |
R. S. Cantrell, C. Cosner, W. F. Fagan, Competitive reversals inside ecological reserves: the role of external habitat degradation, J. Mathe. Biol., 37 (1998), 491-533. doi: 10.1007/s002850050139
|
| [27] |
K. J. Haynes, J. T. Cronin, Matrix composition affects the spatial ecology of a prairie planthopper, Ecology, 84 (2003), 2856-2866. doi: 10.1890/02-0611
|
| [28] | N. Fonseka, J. Goddard Ⅱ, Q. Morris, R. Shivaji, B. Son, On the effects of the exterior matrix hostility and a u-shaped density dependent dispersal on a diffusive logistic growth model, Discrete Continuous Dyn. Syst. Ser. B, Accepted. |
| [29] | J. Goddard Ⅱ, Q. Morris, C. Payne, R. Shivaji, A diffusive logistic equation with u-shaped density dependent dispersal on the boundary, Topol. Methods Nonlinear Anal., 53 (2019), 335-349. |
| [30] |
J. T. Cronin, J. Goddard Ⅱ, R. Shivaji, Effects of patch matrix-composition and individual movement response on population persistence at the patch-level, Bull. Math. Biol., 81 (2019), 3933-3975. doi: 10.1007/s11538-019-00634-9
|
| [31] | J. T. Cronin, N. Fonseka, J. Goddard Ii, J. Leonard, R. Shivaji, Modeling the effects of density dependent emigration, weak allee effects, and matrix hostility on patch-level population persistence, Math. Biosci. Eng., 17 (2019), 1718-1742. |
| [32] |
J. T. Cronin, From population sources to sieves: the matrix alters host-parasitoid source-sink structure, Ecology, 88 (2007), 2966-2976. doi: 10.1890/07-0070.1
|
| [33] |
R. Cantrell, C. Cosner, Y. Lou, Multiple reversals of competitive dominance in ecological reserves via external habitat degradation, J. Dyn. Differ. Equ., 16 (2004), 973-1010. doi: 10.1007/s10884-004-7831-y
|
| [34] |
S. A. Levin, Dispersion and population interactions, Am. Nat., 108 (1974), 207-228. doi: 10.1086/282900
|
| [35] |
S. A. Levin, The role of theoretical ecology in the description and understanding of populations in heterogeneous environments, Am. Zool., 21 (1981), 865-875. doi: 10.1093/icb/21.4.865
|
| [36] | P. C. Fife, Mathematical aspects of reacting and diffusing systems, vol. 28 of Lecture Notes in Biomathematics, Springer-Verlag, 1979. |
| [37] | J. D. Murray, Mathematical Biology. Ⅱ, vol. 18 of Interdisciplinary Applied Mathematics, 3rd edition, Springer-Verlag, New York, 2003. |
| [38] | A. Okubo, Diffusion and ecological problems: mathematical models, vol. 10 of Biomathematics, Springer, Berlin, 1980. |
| [39] | R. S. Cantrell, C. Cosner, Spatial Ecology via Reaction-Diffusion Equations, Mathematical and Computational Biology, Wiley, Chichester, 2003. |
| [40] |
E. E. Holmes, M. A. Lewis, R. R. V. Banks, Partial differential equations in ecology: spatial interactions and population dynamics, Ecology, 75 (1994), 17-29. doi: 10.2307/1939378
|
| [41] | G. A. Maciel, F. Lutscher, H. R. Associate Editor: Sean, D. Editor: Troy, How individual movement response to habitat edges affects population persistence and spatial spread, Am. Nat., 182 (2013), 42-52. |
| [42] |
O. Ovaskainen, Habitat-specific movement parameters estimated using mark-recapture data and a diffusion model, Ecology, 85 (2004), 242-257. doi: 10.1890/02-0706
|
| [43] |
O. Ovaskainen, S. J. Cornell, Biased movement at a boundary and conditional occupancy times for diffusion processes, J. Appl. Probab., 40 (2003), 557-580. doi: 10.1239/jap/1059060888
|
| [44] |
R. S. Cantrell, C. Cosner, On the effects of nonlinear boundary conditions in diffusive logistic equations on bounded domains, J. Differ. Equ., 231 (2006), 768-804. doi: 10.1016/j.jde.2006.08.018
|
| [45] |
R. S. Cantrell, C. Cosner, Density dependent behavior at habitat boundaries and the allee effect, Bull. Math. Biol., 69 (2007), 2339-2360. doi: 10.1007/s11538-007-9222-0
|
| [46] | C. V. Pao, Nonlinear parabolic and elliptic equations, Plenum Press, New York, 1992. |
| [47] |
S. Robinson, M. A. Rivas, Eigencurves for linear elliptic equations, ESAIM Control Optim. Calc. Var., 25 (2019), 45. doi: 10.1051/cocv/2018039
|
| [48] | J. Goddard Ⅱ, Q. Morris, S. Robinson, R. Shivaji, An exact bifurcation diagram for a reaction diffusion equation arising in population dynamics, Bound. Value Probl., 170 (2018), 1-17. |
| [49] | J. T. Cronin, E. Cosgrove, J. Goddard Ⅱ, E. Lindsey, R. Shivaji, Modeling trait-mediated dispersal and habitat fragmentation in two interacting populations, Preprint. |
| [50] |
A. F. G. Dixon, B. K. Agarwala, Ladybird-induced life-history changes in aphids, Proc. Royal Soc. B, 266 (1999), 1549-1553. doi: 10.1098/rspb.1999.0814
|
| [51] |
H. N. Comins, M. P. Hassell, R. M. May, The spatial dynamics of host-parasitoid systems, J. Anim. Ecol., 61 (1992), 735-748. doi: 10.2307/5627
|
| [52] |
J. T. Cronin, K. J. Haynes, An invasive plant promotes unstable host-parasitoid patch dynamics, Ecology, 85 (2004), 2772-2782. doi: 10.1890/04-0303
|
| [53] |
A. Hastings, Spatial heterogeneity and the stability of predator-prey systems, Theor. Popul. Biol., 12 (1977), 37-48. doi: 10.1016/0040-5809(77)90034-X
|
| [54] |
C. Huffaker, Experimental studies on predation: dispersion factors and predator-prey oscillations, Hilgardia, 27 (1958), 343-383. doi: 10.3733/hilg.v27n14p343
|
| [55] |
Y. Yawata, O. X. Cordero, F. Menolascina, J.-H. Hehemann, M. F. Polz, R. Stocker, Competition-dispersal tradeoff ecologically differentiates recently speciated marine bacterioplankton populations, Proc. Natl. Acad. Sci. U. S. A., 111 (2014), 5622-5627. doi: 10.1073/pnas.1318943111
|
| [56] |
M. Cadotte, D. Mai, S. Jantz, M. Collins, M. Keele, J. Drake, On testing the competitioncolonization trade-off in a multispecies assemblage, Am. Nat., 168 (2006), 704-709. doi: 10.1086/508296
|
| [57] |
P. Chesson, General theory of competive coexistence in spatially-varying environments, Theor. Popul. Biol., 58 (2000), 211-237. doi: 10.1006/tpbi.2000.1486
|
| [58] |
R. Levins, D. Culver, Regional coexistence of species and competition between rare species, Proc. Natl. Acad. Sci. U. S. A., 68 (1971), 1246-1248. doi: 10.1073/pnas.68.6.1246
|
| [59] | D. Tilman, P. M. Kareiva, Spatial ecology: the role of space in population dynamics and interspecific interactions, vol. 30, Princeton University Press, 1997. |
| [60] |
P. Amarasekare, The role of density-dependent dispersal in source-sink dynamics, J. Theor. Biol., 226 (2004), 159-168. doi: 10.1016/j.jtbi.2003.08.007
|
| [61] |
A. S. Seymour, D. Gutiérrez, D. Jordano, Dispersal of the lycaenid plebejus argus in response to patches of its mutualist ant lasius niger, Oikos, 103 (2003), 162-174. doi: 10.1034/j.1600-0706.2003.12331.x
|
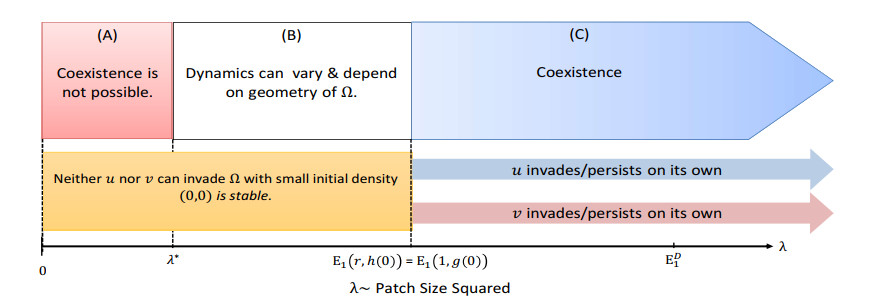
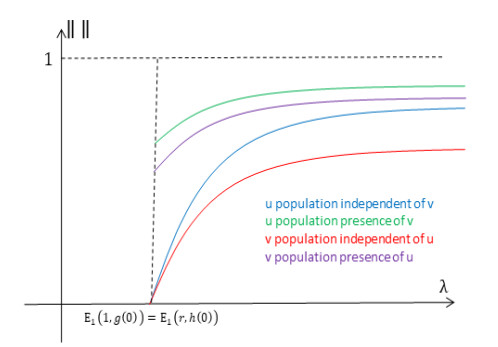
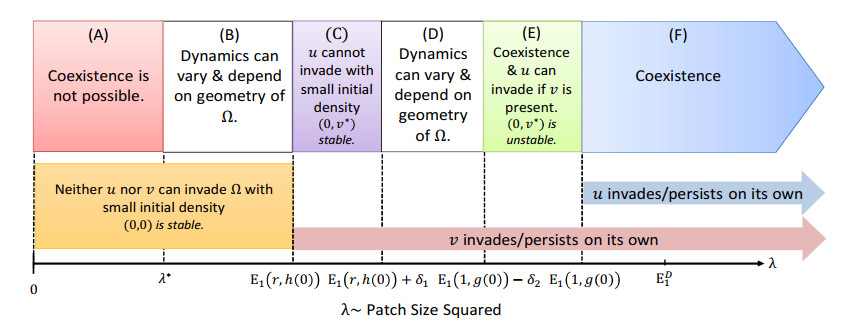
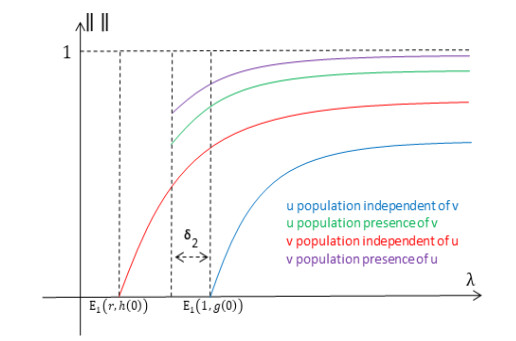

Figures(14) / Tables(1)

James T. Cronin, Jerome Goddard II, Amila Muthunayake, Ratnasingham Shivaji. Modeling the effects of trait-mediated dispersal on coexistence of mutualists[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7838-7861. doi: 10.3934/mbe.2020399







 DownLoad:
DownLoad: