Citation: Abdennasser Chekroun, Mohammed Nor Frioui, Toshikazu Kuniya, Tarik Mohammed Touaoula. Global stability of an age-structured epidemic model with general Lyapunov functional[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1525-1553. doi: 10.3934/mbe.2019073

| [1] | N. Bacaër, A Short History of Mathematical Population Dynamics, Springer-Verlag, London, UK, 2011. |
| [2] | W. O. Kermack and A. G. McKendrick, Contributions to the mathematical theory of epidemics - I, Proc. R. Soc., 115 (1927), 700–721. |
| [3] | O. Diekmann and J. A. P. Heesterbeek, Mathematical epidemiology of infectious diseases: Model building, analysis and interpretation, Wiley, Chichester, UK, 2000. |
| [4] | H. Inaba, Age-Structured Population Dynamics in Demography and Epidemiology, Springer, Singapore, 2017. |
| [5] | P. Magal, C. C. McCluskey and G. F. Webb, Lyapunov functional and global asymptotic stability for an infection-age model, Appl. Anal., 89 (2010), 1109–1140. |
| [6] | Y. Chen, S. Zou and J. Yang, Global analysis of an SIR epidemic model with infection age and saturated incidence, Nonlinear Anal. RWA, 30 (2016), 16–31. |
| [7] | L. Liu, J. Wang and X. Liu, Global stability of an SEIR epidemic model with age-dependent latency and relapse, Nonlinear Anal. RWA, 24 (2015), 18–35. |
| [8] | C. C. McCluskey, Global stability for an SEI epidemiological model with continuous age-structure in the exposed and infectious classes, Math. Biosci. Eng., 9 (2012), 819–841. |
| [9] | C. Vargas-De-Leon, Global stability properties of age-dependent epidemic models with varying rates of recurrence, Math. Meth. Appl. Sci., 39 (2016), 2057–2064. |
| [10] | J. Yang, Y. Chen and T. Kuniya, Threshold dynamics of an age-structured epidemic model with relapse and nonlinear incidence, IMA J. Appl. Math., 82 (2017), 629–655. |
| [11] | A. Chekroun and T. Kuniya, Stability and existence results for a time-delayed nonlocal model of hematopoietic stem cells dynamics, J. Math. Anal. Appl., 463 (2018), 1147–1168. |
| [12] | K. Hattaf and Y. Yang, Global dynamics of an age-structured viral infection model with general incidence function and absorption, Int. J. Biomath., 11 (2018). 1850065 |
| [13] | G. ur Rahman, R. P. Agarwal, L. Liu and A. Khan, Threshold dynamics and optimal control of an age-structured giving up smoking model, Nonlinear Anal. RWA, 43 (2018), 96–120. |
| [14] | V. Capasso and G. Serio, A generalization of the Kermack-McKendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43–61. |
| [15] | S. Djilali, T. M. Touaoula and S. E. H. Miri, A heroin epidemic model: very general non linear incidence, treat-age, and global stability, Acta Applicandae Mathematicae, 152 (2017), 171–194. |
| [16] | Z. Feng and H. R. Thieme, Endemic models with arbitrarily distributed periods of infection I: fundamental properties of the model, SIAM J. Appl. Math., 61 (2000), 803–833. |
| [17] | Z. Feng and H. R. Thieme, Endemic models with arbitrarily distributed periods of infection II: fast disease dynamics and permanent recovery, SIAM J. Appl. Math., 61 (2000), 983–1012. |
| [18] | M. N. Frioui, S. E. Miri and T. M. Touaoula, Unified Lyapunov functional for an age-structured virus model with very general nonlinear infection response, J. Appl. Math. Comput., 58 (2018), 47–73. |
| [19] | K. Hattaf, A. A. Lashari, Y. Louartassi and N. Yousfi, A delayed SIR epidemic model with general incidence rate, Electron. J. Qual. Theo. Diff. Equ., 3 (2013), 1–9. |
| [20] | G. Huang and Y. Takeuchi, Global analysis on delay epidemiological dynamic models with nonlinear incidence, J. Math. Biol., 63 (2011), 125–139. |
| [21] | G. Huang, Y. Takeuchi, W. Ma and D. Wei, Global stability for delay SIR and SEIR epidemic models with nonlinear incidence rate, Bull. Math. Biol., 72 (2010), 1192–1207. |
| [22] | A. Korobeinikov, Lyapunov functions and global stability for SIR and SIRS epidemiological models with non-linear transmission, Bull. Math. Biol., 68 (2006), 615–626. |
| [23] | A. Korobeinikov, Global properties of infectious disease models with nonlinear incidence, Bull. Math. Biol., 69 (2007), 1871-1886. |
| [24] | A. Korobeinikov and P. K. Maini, Nonlinear incidence and stability of infectious disease models, Math. Med. Biol., 22 (2005), 113–128. |
| [25] | S. Liu, S.Wang and L.Wang, Global dynamics of delay epidemic models with nonlinear incidence rate and relapse, Nonlinear Anal. RWA, 12 (2011), 119–127. |
| [26] | J.Wang, J. Pang and X. Liu, Modelling diseases with relapse and nonlinear incidence of infection: a multi-group epidemic model, J. Biol. Dyn., 8 (2014), 99–116. |
| [27] | S. Bentout and T. M. Touaoula, Global analysis of an infection age model with a class of nonlinear incidence rates, J. Math. Anal. Appl., 434 (2016), 1211–1239. |
| [28] | P. van den Driessche and X. Zou, Modeling relapse in infectious diseases, Math. Biosci., 207 (2007), 89–103. |
| [29] | B. Fang, X. Z. Li, M. Martcheva and L. M. Cai, Global asymptotic properties of a heroin epidemic model with treat-age, Appl. Math. Comput., 263 (2015), 315–331. |
| [30] | G. Mulone and B. Straughan, A note on heroin epidemics, Math. Biosci., 218 (2009), 138–141. |
| [31] | Y. Muroya, H. Li and T. Kuniya, Complete global analysis of an SIRS epidemic model with grated cure and incomplete recovery rates, J. Math. Anal. Appl., 410 (2014), 719–732. |
| [32] | E. White and C. Comiskey, Heroin epidemics, treatment and ODE modelling, Math. Biosci. 208 (2007), 312–324. |
| [33] | I. M. Wangari and L. Stone, Analysis of a heroin epidemic model with saturated treatment function, J. Appl. Math., 2017 (2017), Article ID 1953036. |
| [34] | P. Munz, I. Hudea, J. Imad and R. J. Smith, When zombies attack!: mathematical modelling of an outbreak of zombie infection, in Infectious Disease Modelling Research Progress (eds. J.M. Tchuenche and C. Chiyaka), New York, Nova Science Publishers, (2009), 133–150. |
| [35] | J. K. Hale and S. M. Verduyn Lunel, Introduction to Functional Differential Equations, V. 99, AMS, 1993. |
| [36] | B. Balachandran, T. Kalmar-Nagy and D. E. Gilsin, Delay Differential Equations: Recent Advances and New Direction, Springer-verlag, 2009. |
| [37] | H. L. Smith and H. R. Thieme, Dynamical Systems and Population Persistence, Graduate Studies in Mathematics V. 118, AMS, 2011. |
| [38] | H. R. Thieme, Mathematics in Population Biology, Princeton University Press, Princeton 2003. |
| [39] | B. Perthame, Transport Equations in Biology, Birkhäuser, Berlin, 2007. |
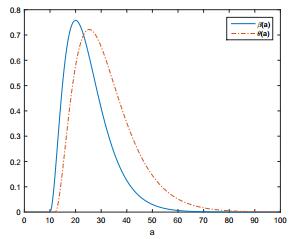
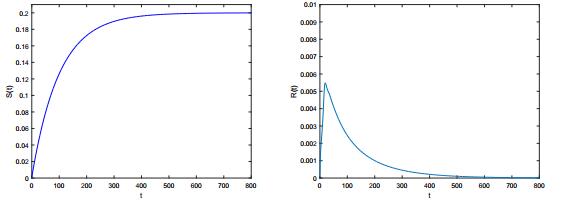
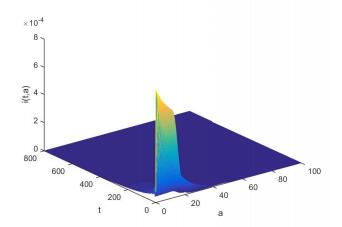
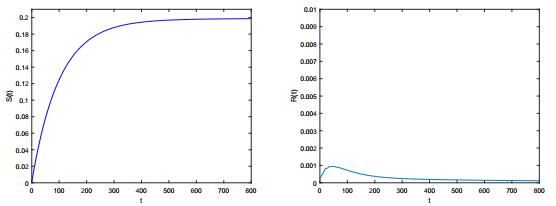
Figures(6)

Abdennasser Chekroun, Mohammed Nor Frioui, Toshikazu Kuniya, Tarik Mohammed Touaoula. Global stability of an age-structured epidemic model with general Lyapunov functional[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1525-1553. doi: 10.3934/mbe.2019073







 DownLoad:
DownLoad: