Citation: Chang-Yuan Cheng, Kuang-Hui Lin, Chih-Wen Shih. Coexistence and extinction for two competing species in patchy environments[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 909-946. doi: 10.3934/mbe.2019043

| [1] | E. Braveman and Md. Kamrujjamam, Lotka systems with directed dispersal dynamics: Competition and influence of diffusion strategies, Math. Biosci., 279 (2016), 1–12. |
| [2] | R. S. Cantrell and C. Cosner, Spatial ecology via reaction-diffusion equations, Series in Mathematical and Computational Biology, John Wiley and Sons, Chichester, UK, 2003. |
| [3] | R. S. Cantrell, C. Cosner and Y. Lou, Multiple reversals of competitive dominance in ecological reserves via external habitat degradation, J. Dyn. Diff. Eqs., 16 (2004), 973–1010. |
| [4] | R. S. Cantrell, C. Cosner, D. L. DeAngelis and V. Padron, The ideal free distribution as an evolutionarily stable strategy, J. Biol. Dyn., 1 (2007), 249–271. |
| [5] | R. S. Cantrell, C. Cosner and Y. Lou, Evolutionary stability of ideal free dispersal strategies in patchy environments, J. Math. Biol., 65 (2012), 943–965. |
| [6] | X. Chen, K. Y. Lam and Y. Lou, Dynamics of a reaction-diffusion-advection model for two competing species, Discret. Contin. Dyn. Syst., 32 (2012), 3841–3859. |
| [7] | J. Clobert, E. Danchin, A. A. Dhondt and J. D. Nichols, Dispersal, Oxford University Press, 2001. |
| [8] | R. Cressman and V. Krivan, Two-patch population models with adaptive dispersal the effects of varying dispersal speeds, J. Math. Biol., 67 (2013), 329–358. |
| [9] | J. Dockery, V. Hutson, K. Mischaikow and M. Pernarowski, The evolution of slow dispersal rates: a reaction diffusion model, J. Math. Biol., 37 (1998), 61–83. |
| [10] | G. F. Gause, The Struggle for Existence, Williams Wilkins, Baltimore, MD, 1934. |
| [11] | S. A. Gourley and Y. Kuang, Two-species competition with high dispersal: the winning strategy, Math. Biosci. Eng., 2 (2005), 345–362. |
| [12] | A. Hastings, Can spatial variation alone lead to selection for dispersal?, Theor. Popul. Biol., 24 (1983), 244–251. |
| [13] | X. Q. He and W. M. Ni, The effects of diffusion and spatial variation in LotkaVolterra competitiondi ffusion system I: Heterogeneity vs. homogeneity, J. Differ. Eqs., 254 (2013), 528–546. |
| [14] | X. Q. He and W. M. Ni, The effects of diffusion and spatial variation in LotkaVolterra competitiondi ffusion system II: The general case, J. Differ. Eqs., 254 (2013), 4088–4108. |
| [15] | X. Q. He and W. M. Ni, Global dynamics of the Lotka-Volterra competition-diffusion system: diffusion and spatial heterogeneity I, Commun. Pure Appl. Math., LXIX (2016), 981–1014. |
| [16] | R. D. Holt, Population dynamics in two-patch environments some anomalous consequences of an optimal habitat distribution, Theor. Popul. Biol., 28 (1985), 181–208. |
| [17] | S. B. Hsu, H. Smith and P. Waltman, Competitive exclusion and coexistence for competitive systems on ordered Banach spaces, Trans. Amer. Math. Soc., 348 (1996), 4083–4094. |
| [18] | V. Hutson, J. Lopez-Gomez, K. Mischaikow and G. Vickers, Limit behaviour for a competing species problem with diffusion, in Dynamical Systems and Applications, in World Sci. Ser. Appl. Anal. 4, World Scientific, River Edge, NJ, (1995), 343–358. |
| [19] | V. Hutson, Y. Lou, K. Mischaikow and P. Poláčik, Competing species near a degenerate limit, SIAM. J. Math. Anal., 35 (2003), 453–491. |
| [20] | V. Hutson, S. Martinez, K. Mischaikow and G. T. Vickers, The evolution of dispersal, J. Math. Biol., 47 (2003), 483–517. |
| [21] | J. Jiang and X. Liang, Competitive systems with migration and the Poincaré-Bendixson theorem for a 4-dimensional case, Quar. Appl. Math., LXIV (2006), 483–498. |
| [22] | Y. Kuang and Y. Takeuchi, Predator-prey dynamics in models of prey dispersal in two-patch environments, Math. Biosci., 120 (1994), 77–98. |
| [23] | K. Y. Lam and W. M. Ni, Uniqueness and complete dynamics in heterogeneous competitiondi ffusion systems, SIAM J. Appl. Math., 72 (2012), 1695–1712. |
| [24] | K. H. Lin, Y. Lou, C. W. Shih and T. H. Tsai, Global dynamics for two-species competition in patchy environment, Math. Biosci. Eng., 11 (2014), 947–970. |
| [25] | Y. Lou, S. Martinez and P. Polacik, Loops and branches of coexistence states in a Lotka-Volterra competition model, J. Differ. Eqs., 230 (2006), 720–742. |
| [26] | A. Okubo and S. A. Levin, Diffusion and ecological problems: modern perspectives, second ed., Interdisciplinary Applied Mathematics, 14, Springer, New York, 2001. |
| [27] | H. L. Smith, Competing subcommunities of mutualists and a generalized Kamke theorem, SIAM J. Appl. Math., 46 (1986), 856–874. |
| [28] | H. L. Smith, Monotone Dynamical Systems: an introduction to the theory of competitive and cooperative systems, Math. Surveys and Monographs, Amer. Math. Soc., 41, 1995. |
| [29] | Y. Takeuchi, Diffusion-mediated persistence in two-species competition Lotka-Volterra model, Math. Biosci., 95 (1989), 65–83. |
| [30] | Y. Takeuchi and Z. Lu, Permanence and global stability for competitive Lotka-Volterra diffusion systems, Nonlinear Anal. TMA, 24 (1995), 91–104. |
| [31] | Y. Takeuchi, Global dynamical properties of Lotka-Volterra systems, River Edge, NJ 07661, World Scientic, 1996. |
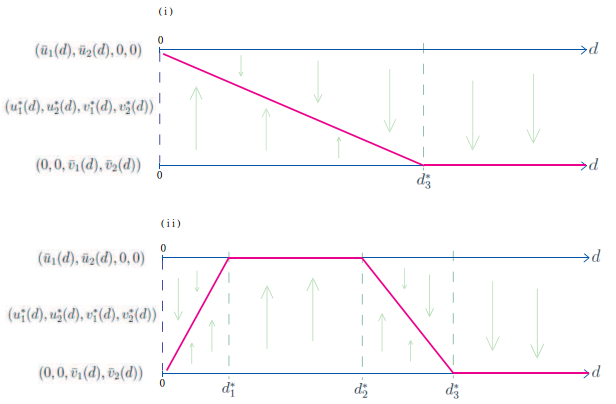
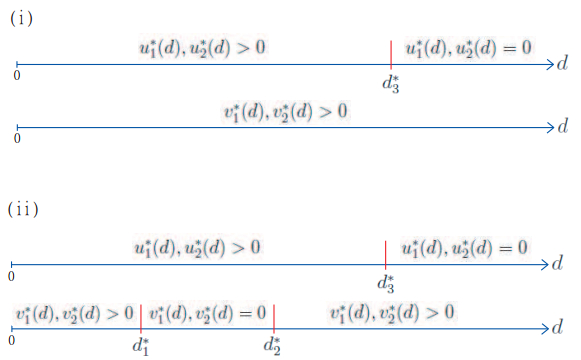
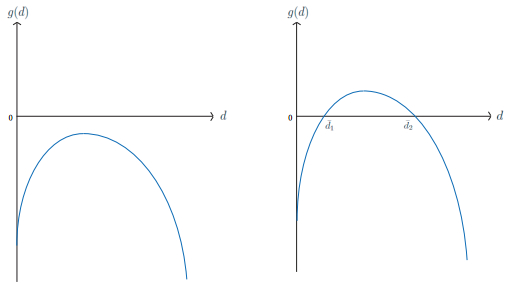

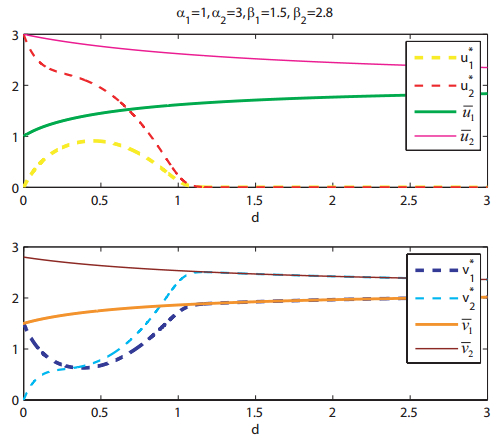
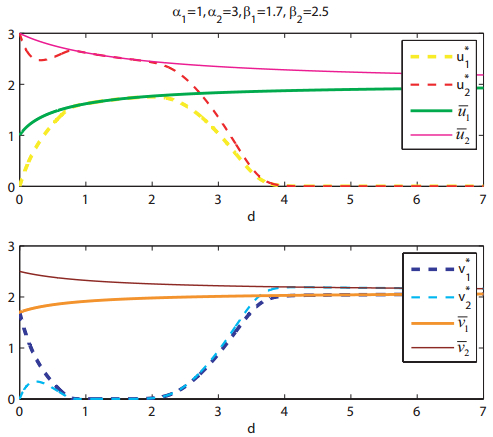
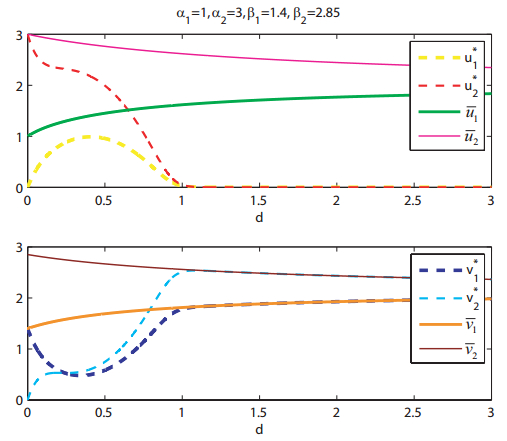
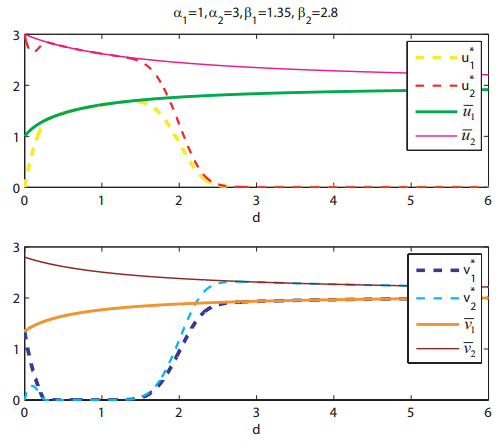
Figures(8)

Chang-Yuan Cheng, Kuang-Hui Lin, Chih-Wen Shih. Coexistence and extinction for two competing species in patchy environments[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 909-946. doi: 10.3934/mbe.2019043







 DownLoad:
DownLoad: