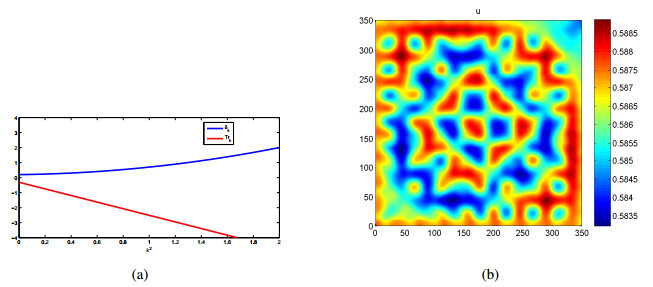
We introduce a diffusive predator-prey system with the double Allee effect, focusing on the stability and sufficient conditions for the coexistence of prey and predator. Subsequently, we derived the amplitude equation and explore secondary-order dynamic properties using methods such as Taylor series expansion and multiscaling. The novel approach outlined above provides a precise means to thoroughly analyze the predator-prey model. Through this analysis, we demonstrated that the inclusion of the Allee effect and diffusion leads to the system exhibiting more intricate dynamic behaviors compared to systems lacking these factors. On one hand, in the diffusive system without the Allee effect, the pattern formation regarding the distribution of species was relatively scattered, whereas in the diffusive system with the Allee effect, it is more intensive. On the other hand, the system with the Allee effect transitioned from unstable to stable when the diffusion parameter in prey increased, and the aggregation degree of pattern formation in the system with the Allee effect was higher than in the system without it. These findings highlight the significant roles played by the Allee effect and diffusion in determining the dynamic behaviors of prey and predator within the system.
Citation: Lingling Li, Xuechen Li. The spatiotemporal dynamics of a diffusive predator-prey model with double Allee effect[J]. AIMS Mathematics, 2024, 9(10): 26902-26915. doi: 10.3934/math.20241309
We introduce a diffusive predator-prey system with the double Allee effect, focusing on the stability and sufficient conditions for the coexistence of prey and predator. Subsequently, we derived the amplitude equation and explore secondary-order dynamic properties using methods such as Taylor series expansion and multiscaling. The novel approach outlined above provides a precise means to thoroughly analyze the predator-prey model. Through this analysis, we demonstrated that the inclusion of the Allee effect and diffusion leads to the system exhibiting more intricate dynamic behaviors compared to systems lacking these factors. On one hand, in the diffusive system without the Allee effect, the pattern formation regarding the distribution of species was relatively scattered, whereas in the diffusive system with the Allee effect, it is more intensive. On the other hand, the system with the Allee effect transitioned from unstable to stable when the diffusion parameter in prey increased, and the aggregation degree of pattern formation in the system with the Allee effect was higher than in the system without it. These findings highlight the significant roles played by the Allee effect and diffusion in determining the dynamic behaviors of prey and predator within the system.

| [1] | A. J. Lotka, A Natural Population Norm I and II, Washington Academy of Sciences, 1913. |
| [2] |
V. Volterra, Fluctuations in the abundance of a species considered mathematically, Nature, 118 (1926), 558–560. http://dx.doi.org/10.1038/118558a0 doi: 10.1038/118558a0

|
| [3] | W. C. Allee, Animal Aggregations, University of Chicago Press, 1931. |
| [4] |
G. Q. Sun, Mathematical modeling of population dynamics with Allee effect, Nonlinear Dynam., 85 (2016), 1–12. http://dx.doi.org/10.1007/s11071-016-2671-y doi: 10.1007/s11071-016-2671-y
|
| [5] |
M. Groom, Allee effects limit population viability of an annual plant, Am. Nat., 151 (1998), 487–496. http://dx.doi.org/10.2307/2463323 doi: 10.2307/2463323
|
| [6] | F. Courchamp, J. Berec, J. Gascoigne, Allee effects in ecology and conservation, Oxford University Press, 2008. |
| [7] |
P. A. Stephens, W. J. Sutherland, R. P. Freckleton, What is the Allee effect, Nordic Society Oikos, 87 (1999), 185–190. http://dx.doi.org/10.1007/s003960050147 doi: 10.1007/s003960050147
|
| [8] |
Y. N. Zeng, P. Yu, Complex dynamics of predator-prey systems with Allee effect, Int. J. Bifurcat. Chaos, 32 (2022), 2250203. http://dx.doi.org/10.1142/S0218127422502030 doi: 10.1142/S0218127422502030
|
| [9] |
J. D. Flores, E. Gonzalez-Olivares, Dynamics of a predator-prey model with Allee effect on prey and ratio-dependent functional response, Ecol. Complex., 18 (2014), 59–66. http://dx.doi.org/10.1016/j.ecocom.2014.02.005 doi: 10.1016/j.ecocom.2014.02.005
|
| [10] |
H. Molla, S. Sarwardi, S. R. Smith, M. Haque, Dynamics of adding variable prey refuge and an Allee effect to a predator-prey model, Alex. Eng. J., 61 (2022), 4175–4188. http://dx.doi.org/10.1016/j.aej.2021.09.039 doi: 10.1016/j.aej.2021.09.039
|
| [11] |
A. J. Terry, Prey resurgence from mortality events in predator-prey models, Nonlinear Anal-Real, 14 (2013), 2180–2203. http://dx.doi.org/10.1016/j.nonrwa.2013.04.006 doi: 10.1016/j.nonrwa.2013.04.006
|
| [12] |
A. J. Terry, A predator-prey model with generic birth and death rates for the predator, Math. Biosci., 248 (2014), 57–66. http://dx.doi.org/10.1016/j.mbs.2013.12.002 doi: 10.1016/j.mbs.2013.12.002
|
| [13] |
A. F. Rozenfeld, C. J. Tessone, E. Albano, H. S. Wio, On the influence of noise on the critical and oscillatory behavior of a predator-prey model: coherent stochastic resonance at the proper frequency of the system, Phys. Lett. A, 280 (2001), 45–52. http://dx.doi.org/10.1016/S0375-9601(01)00033-0 doi: 10.1016/S0375-9601(01)00033-0
|
| [14] |
L. L. Li, J. W. Shen, Bifurcations and dynamics of a predator-prey model with double Allee effects and time delays, Int. J. Bifurcat. Chaos, 28 (2018), 1850135. http://dx.doi.org/10.1142/s0218127418501353 doi: 10.1142/s0218127418501353
|
| [15] |
Z. P. Ma, Spatiotemporal dynamics of a diffusive Leslie-Gower prey-predator model with strong Allee effect, Nonlinear Anal-Real, 50 (2019), 651–674. http://dx.doi.org/10.1016/j.nonrwa.2019.06.008 doi: 10.1016/j.nonrwa.2019.06.008
|
| [16] |
Q. Q. zheng, J. W. Shen, V. Pandey, X. P. Yuan, L. N. Guan, Pattern selection mechanism from the equilibrium point and limit cycle, Chaos, 34 (2024), 023124. http://dx.doi.org/10.1063/5.0187810 doi: 10.1063/5.0187810
|
| [17] |
Q. Q. Zheng, J. W. Shen, V. Pandey, Y. M. Zhao, L. N. Guan, Spatiotemporal dynamics of periodic waves in SIR model with driving factors, New J. Phys., 25 (2023), 063028. http://dx.doi.org/10.1088/1367-2630/acdb91 doi: 10.1088/1367-2630/acdb91
|
| [18] |
Q. Q. Zheng, J. W. Shen, Y. Xu, Turing instability in the reaction-diffusion network, Phys. Review E, 102 (2020), 062215. http://dx.doi.org/10.1103/PhysRevE.102.062215 doi: 10.1103/PhysRevE.102.062215
|
| [19] |
Q. Hu, J. W. Shen, Turing instability of the modified reaction-diffusion Holling-Tanner model in random network, Int. J. Bifurcat. Chaos, 32 (2022), 1–18. http://dx.doi.org/10.1142/S0218127422500493 doi: 10.1142/S0218127422500493
|
| [20] |
A. M. Turing, The chemical basis of morphogenesis, B. Math. Biol., 52 (1990), 153–197. http://dx.doi.org/10.1007/BF02459572 doi: 10.1007/BF02459572
|
| [21] | Q. Ouyang, Introduction to nonlinear science and pattern dynamics, Peking university press, 2010. |
| [22] |
X. Z. Fan, F. Zhou, Y. Li, Stationary pattern and Hopf bifurcation of a diffusive predator-prey model, Appl. Anal., 102 (2023), 2141–2159. http://dx.doi.org/10.1080/00036811.2021.2021186 doi: 10.1080/00036811.2021.2021186
|
| [23] |
G. Q. Sun, M. Jusup, Z. Jin, Y. Wang, Z. Wang, Pattern transitions in spatial epidemics: Mechanisms and emergent properties, Phys. Life Rev., 19 (2016), 43–73. http://dx.doi.org/10.1016/j.plrev.2016.08.002 doi: 10.1016/j.plrev.2016.08.002
|
| [24] |
Q. Q. Zheng, J. W. Shen, Pandey, V, L. A. Guan, Y. T. Guo, Turing instability in a network-organized epidemic model with delay, Chaos, Soliton. Fract., 168 (2023), 113205. http://dx.doi.org/10.1016/j.chaos.2023.113205 doi: 10.1016/j.chaos.2023.113205
|
| [25] |
R. H. William, E. G. Adriana, M. B. William, E. K. Leah, A mathematical model of GTPase pattern formation during single-cell wound repair, Interface Focus, 6 (2016), 20160032. http://dx.doi.org/10.1098/rsfs.2016.0032 doi: 10.1098/rsfs.2016.0032
|
| [26] | Q. Ouyang, Patterns formation in reaction diffusion systems, Shanghai Sci.-Tech Education Publishing House, 2000. |





Figures(6)

Lingling Li, Xuechen Li. The spatiotemporal dynamics of a diffusive predator-prey model with double Allee effect[J]. AIMS Mathematics, 2024, 9(10): 26902-26915. doi: 10.3934/math.20241309







 DownLoad:
DownLoad: