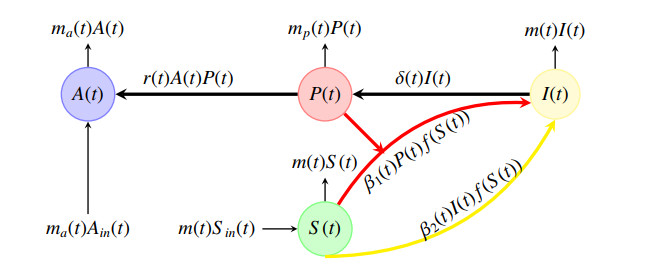
The chikungunya virus (CHIKV) infects macrophages and adherent cells and it can be transmitted via a direct contact with the virus or with an already infected cell. Thus, the CHIKV infection can have two routes. Furthermore, it can exhibit seasonal peak periods. Thus, in this paper, we consider a dynamical system model of the CHIKV dynamics under the conditions of a seasonal environment with a general incidence rate and two routes of infection. In the first step, we studied the autonomous system by investigating the global stability of the steady states with respect to the basic reproduction number. In the second step, we establish the existence, uniqueness, positivity and boundedness of a periodic orbit for the non-autonomous system. We show that the global dynamics are determined by using the basic reproduction number denoted by $ \mathcal{R}_0 $ and they are calculated using the spectral radius of an integral operator. We show the global stability of the disease-free periodic solution if $ \mathcal{R}_0 < 1 $ and we also show the persistence of the disease if $ \mathcal{R}_0 > 1 $ where the trajectories converge to a limit cycle. Finally, we display some numerical investigations supporting the theoretical findings.
Citation: Miled El Hajji. Periodic solutions for chikungunya virus dynamics in a seasonal environment with a general incidence rate[J]. AIMS Mathematics, 2023, 8(10): 24888-24913. doi: 10.3934/math.20231269
The chikungunya virus (CHIKV) infects macrophages and adherent cells and it can be transmitted via a direct contact with the virus or with an already infected cell. Thus, the CHIKV infection can have two routes. Furthermore, it can exhibit seasonal peak periods. Thus, in this paper, we consider a dynamical system model of the CHIKV dynamics under the conditions of a seasonal environment with a general incidence rate and two routes of infection. In the first step, we studied the autonomous system by investigating the global stability of the steady states with respect to the basic reproduction number. In the second step, we establish the existence, uniqueness, positivity and boundedness of a periodic orbit for the non-autonomous system. We show that the global dynamics are determined by using the basic reproduction number denoted by $ \mathcal{R}_0 $ and they are calculated using the spectral radius of an integral operator. We show the global stability of the disease-free periodic solution if $ \mathcal{R}_0 < 1 $ and we also show the persistence of the disease if $ \mathcal{R}_0 > 1 $ where the trajectories converge to a limit cycle. Finally, we display some numerical investigations supporting the theoretical findings.

| [1] | H. W. Hethcote, Three basic epidemiological models, In: S. A. Levin, T. G. Hallam, L. J. Gross, Applied mathematical ecology, Biomathematics, Springer, Berlin, Heidelberg, 18 (1989), 119–144. https://doi.org/10.1007/978-3-642-61317-3_5 |
| [2] |
A. Alshehri, M. El Hajji, Mathematical study for Zika virus transmission with general incidence rate, AIMS Math., 7 (2022), 7117–7142. https://doi.org/10.3934/math.2022397 doi: 10.3934/math.2022397

|
| [3] |
M. El Hajji, A. H. Albargi, A mathematical investigation of an "SVEIR" epidemic model for the measles transmission, Math. Biosci. Eng., 19 (2022), 2853–2875. https://doi.org/10.3934/mbe.2022131 doi: 10.3934/mbe.2022131
|
| [4] |
M. El Hajji, S. Sayari, A. Zaghdani, Mathematical analysis of an "SIR" epidemic model in a continuous reactor–Deterministic and probabilistic approaches, J. Korean Math. Soc., 58 (2021), 45–67. https://doi.org/10.4134/JKMS.j190788 doi: 10.4134/JKMS.j190788
|
| [5] |
S. Alsahafi, S. Woodcock, Mathematical study for chikungunya virus with nonlinear general incidence rate, Mathematics, 9 (2021), 2186. https://doi.org/10.3390/math9182186 doi: 10.3390/math9182186
|
| [6] |
A. Elaiw, T. Alade, S. Alsulami, Analysis of within-host CHIKV dynamics models with general incidence rate, Int. J. Biomath., 11 (2018), 1850062. https://doi.org/10.1142/S1793524518500626 doi: 10.1142/S1793524518500626
|
| [7] | A. Elaiw, S. Almalki, A. Hobiny, Global dynamics of chikungunya virus with two routes of infection, J. Comput. Anal. Appl., 28 (2020), 481–490. |
| [8] |
P. Pinto, M. A. Costa, M. F. M. Gonçalves, A. G. Rodrigues, C. Lisboa, Mpox person-to-person transmission–Where have we got so far? A systematic review, Viruses, 15 (2023), 1074. https://doi.org/10.3390/v15051074 doi: 10.3390/v15051074
|
| [9] |
A. A. Alsolami, M. El Hajji, Mathematical analysis of a bacterial competition in a continuous reactor in the presence of a virus, Mathematics, 11 (2023), 883. https://doi.org/10.3390/math11040883 doi: 10.3390/math11040883
|
| [10] |
M. Al-Raeei, The study of human monkeypox disease in 2022 using the epidemic models: herd immunity and the basic reproduction number case, Ann. Med. Surg. (Lond.), 85 (2023), 316–321. https://doi.org/10.1097/MS9.0000000000000229 doi: 10.1097/MS9.0000000000000229
|
| [11] |
V. Mahmoud, G. Hatem, A. Al-Saleh, D. Ghanem, A. Yassine, S. Awada, Predictors of all-cause mortality in hospitalized Covid-19 patients taking corticosteroids: a multicenter retrospective cross-sectional study, Ann. Med. Surg. (Lond.), 85 (2023), 3386–3395. https://doi.org/10.1097/MS9.0000000000000946 doi: 10.1097/MS9.0000000000000946
|
| [12] |
A. H. Albargi, M. El Hajji, Bacterial competition in the presence of a virus in a chemostat, Mathematics, 11 (2023), 3530. https://doi.org/10.3390/math11163530 doi: 10.3390/math11163530
|
| [13] |
N. Bacaër, M. G. M. Gomes, On the final size of epidemics with seasonality, Bull. Math. Biol., 71 (2009), 1954–1966. https://doi.org/10.1007/s11538-009-9433-7 doi: 10.1007/s11538-009-9433-7
|
| [14] |
J. Ma, Z. Ma, Epidemic threshold conditions for seasonally forced SEIR models, Math. Biosci. Eng., 3 (2006), 161–172. https://doi.org/10.3934/mbe.2006.3.161 doi: 10.3934/mbe.2006.3.161
|
| [15] | S. Guerrero-Flores, O. Osuna, C. V. de Leon, Periodic solutions for seasonal SIQRS models with nonlinear infection terms, Electron. J. Differ. Eq., 2019 (2019), 1–13. |
| [16] |
T. Zhang, Z. Teng, On a nonautonomous SEIRS model in epidemiology, Bull. Math. Biol., 69 (2007), 2537–2559. https://doi.org/10.1007/s11538-007-9231-z doi: 10.1007/s11538-007-9231-z
|
| [17] |
Y. Nakata, T. Kuniya, Global dynamics of a class of SEIRS epidemic models in a periodic environment, J. Math. Anal. Appl., 363 (2010), 230–237. https://doi.org/10.1016/j.jmaa.2009.08.027 doi: 10.1016/j.jmaa.2009.08.027
|
| [18] |
N. Bacaër, S. Guernaoui, The epidemic threshold of vector-borne diseases with seasonality, J. Math. Biol., 53 (2006), 421–436. https://doi.org/10.1007/s00285-006-0015-0 doi: 10.1007/s00285-006-0015-0
|
| [19] |
W. Wang, X. Zhao, Threshold dynamics for compartmental epidemic models in periodic environments, J. Dyn. Diff. Equat., 20 (2008), 699–717. https://doi.org/10.1007/s10884-008-9111-8 doi: 10.1007/s10884-008-9111-8
|
| [20] |
M. El Hajji, D. M. Alshaikh, N. A. Almuallem, Periodic behaviour of an epidemic in a seasonal environment with vaccination, Mathematics, 11 (2023), 2350. https://doi.org/10.3390/math11102350 doi: 10.3390/math11102350
|
| [21] |
M. El Hajji, Modelling and optimal control for chikungunya disease, Theory Biosci., 140 (2021), 27–44. https://doi.org/10.1007/s12064-020-00324-4 doi: 10.1007/s12064-020-00324-4
|
| [22] |
M. El Hajji, A. Zaghdani, S. Sayari, Mathematical analysis and optimal control for chikungunya virus with two routes of infection with nonlinear incidence rate, Int. J. Biomath., 15 (2022), 2150088. https://doi.org/10.1142/S1793524521500881 doi: 10.1142/S1793524521500881
|
| [23] |
A. H. Albargi, M. El Hajji, Mathematical analysis of a two-tiered microbial food-web model for the anaerobic digestion process, Math. Biosci. Eng., 20 (2023), 6591–6611. https://doi.org/10.3934/mbe.2023283 doi: 10.3934/mbe.2023283
|
| [24] |
O. Diekmann, J. A. P. Heesterbeek, J. A. J. Metz, On the definition and the computation of the basic reproduction ratio $\mathcal{R}_0$ in models for infectious diseases in heterogeneous populations, J. Math. Bio., 28 (1990), 365–382. https://doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324
|
| [25] | F. S. Roberts, Graph theory and its applications to problems of society, Philadelphia: Society for Industrial and Applied Mathematics, 1978. https://doi.org/10.1137/1.9781611970401 |
| [26] |
A. Sisk, N. Fefferman, A network theoretic method for the basic reproductive number for infectious diseases, Methods Ecol. Evol., 13 (2022), 2503–2515. https://doi.org/10.1111/2041-210X.13978 doi: 10.1111/2041-210X.13978
|
| [27] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/s0025-5564(02)00108-6 doi: 10.1016/s0025-5564(02)00108-6
|
| [28] |
O. Diekmann, J. Heesterbeek, M. Roberts, The construction of next-generation matrices for compartmental epidemic models, J. R. Soc. Interface, 7 (2010), 873–885. https://doi.org/10.1098/rsif.2009.0386 doi: 10.1098/rsif.2009.0386
|
| [29] | J. LaSalle, The stability of dynamical systems, Philadelphia: Society for Industrial and Applied Mathematics, 1976. https://doi.org/10.1137/1.9781611970432 |
| [30] |
F. Zhang, X. Zhao, A periodic epidemic model in a patchy environment, J. Math. Anal. Appl., 325 (2007), 496–516. https://doi.org/10.1016/j.jmaa.2006.01.085 doi: 10.1016/j.jmaa.2006.01.085
|
| [31] | X. Zhao, Dynamical systems in population biology, CMS Books in Mathematics, Springer Cham, 2003. https://doi.org/10.1007/978-3-319-56433-3 |








Figures(9) / Tables(3)

Miled El Hajji. Periodic solutions for chikungunya virus dynamics in a seasonal environment with a general incidence rate[J]. AIMS Mathematics, 2023, 8(10): 24888-24913. doi: 10.3934/math.20231269







 DownLoad:
DownLoad: