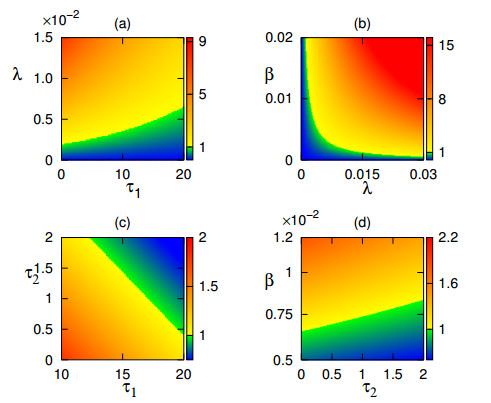
A mathematical model is developed for analysis of the spread of mosaic disease in plants, which account for incubation period and latency that are represented by time delays. Feasibility and stability of different equilibria are studied analytically and numerically. Conditions that determine the type of behavior exhibited by the system are found in terms of various parameters. We have derived the basic reproduction number and identify the conditions resulting in eradication of the disease, as well as those that lead to the emergence of stable oscillations in the population of infected plants, as a result of Hopf bifurcation of the endemic equilibrium. Numerical simulations are performed to verify the analytical results and also to illustrate different dynamical regimes that can be observed in the system. In this research, the stabilizing role of both the time delay has been established i.e. when delay time is large, disease will persist if the infection rate is higher. The results obtained here are useful for plant disease management.
Citation: Fahad Al Basir, Konstantin B. Blyuss, Ezio Venturino. Stability and bifurcation analysis of a multi-delay model for mosaic disease transmission[J]. AIMS Mathematics, 2023, 8(10): 24545-24567. doi: 10.3934/math.20231252
A mathematical model is developed for analysis of the spread of mosaic disease in plants, which account for incubation period and latency that are represented by time delays. Feasibility and stability of different equilibria are studied analytically and numerically. Conditions that determine the type of behavior exhibited by the system are found in terms of various parameters. We have derived the basic reproduction number and identify the conditions resulting in eradication of the disease, as well as those that lead to the emergence of stable oscillations in the population of infected plants, as a result of Hopf bifurcation of the endemic equilibrium. Numerical simulations are performed to verify the analytical results and also to illustrate different dynamical regimes that can be observed in the system. In this research, the stabilizing role of both the time delay has been established i.e. when delay time is large, disease will persist if the infection rate is higher. The results obtained here are useful for plant disease management.

| [1] | D. S. A. Narayana, K. S. Shankarappa, M. R. Govindappa, H. A. Prameela, M. R. G. Rao, K. T. Rangaswamy, Natural occurrence of Jatropha mosaic virus disease in India, Curr. Sci., 91 (2006), 584–586. |
| [2] | O. J. Alabi, P. L. Kumar, R. A. Naidu, Cassava mosaic disease: A curse to food security in sub-Saharan Africa, APSnet Features, 2011, 1–17. |
| [3] | J. E. Duffus, Whitefly transmission of plant viruses, In: Current topics in vector research, New York: Springer, 1987, 73–91. https://doi.org/10.1007/978-1-4612-4712-8_3 |
| [4] |
S. Q. Gao, J. Qu, N. H. Chua, J. Ye, A new strain of Indian cassava mosaic virus causes a mosaic disease in the biodiesel crop Jatropha curcas, Arch. Virol., 155 (2010), 607–612. https://doi.org/10.1007/s00705-010-0625-0 doi: 10.1007/s00705-010-0625-0

|
| [5] | J. P. Tewari, H. D. Dwivedi, P. Madhvi, S. K. Srivastava, Incidence of a mosaic disease in Jatropha curcas L. from eastern Uttar Pradesh, Curr. Sci., 93 (2007), 1048–1049. |
| [6] |
B. D. Kashina, M. D. Alegbejo, O. O. Banwo, S. L. Nielsen, M. Nicolaisen, Molecular identification of a new begomovirus associated with mosaic disease of Jatropha curcas L. in Nigeria, Arch. Virol., 158 (2013), 511–514. https://doi.org/10.1007/s00705-012-1512-7 doi: 10.1007/s00705-012-1512-7
|
| [7] |
H. Czosnek, A. Hariton-Shalev, I. Sobol, R. Gorovits, M. Ghanim, The incredible journey of begomoviruses in their whitefly vector, Viruses, 9 (2017), 273. https://doi.org/10.3390/v9100273 doi: 10.3390/v9100273
|
| [8] |
A. S. Costa, Whitefly-transmitted plant diseases, Annu. Rev. Phytopathol., 14 (1976), 429–449. https://doi.org/10.1146/annurev.py.14.090176.002241 doi: 10.1146/annurev.py.14.090176.002241
|
| [9] |
M. Leclerc, T. Doré, C. A. Gilligan, P. Lucas, J. A. N. Filipe, Estimating the delay between host infection and disease (incubation period) and assessing its significance to the epidemiology of plant diseases, PLoS One, 9 (2014), e86568. https://doi.org/10.1371/journal.pone.0086568 doi: 10.1371/journal.pone.0086568
|
| [10] |
A. van Maanen, X. M. Xu, Modelling plant disease epidemics, Eur. J. Plant Path., 109 (2003), 669–682. https://doi.org/10.1023/A:1026018005613 doi: 10.1023/A:1026018005613
|
| [11] |
B. Buonomo, M. Cerasuolo, The effect of time delay in plant-pathogen interactions with host demography, Math. Biosci. Eng., 12 (2015), 473–490. https://doi.org/10.3934/mbe.2015.12.473 doi: 10.3934/mbe.2015.12.473
|
| [12] | T. Erneux, Applied delay differential equations, New York: Springer, 2009. https://doi.org/10.1007/978-0-387-74372-1 |
| [13] | Y. Kuang, Delay differential equations with applications in population dynamics, New York: Academic Press, 1993. |
| [14] | J. E. Duffus, Whitefly transmission of plant viruses, In: Current topics in vector research, New York: Springer, (1987) 73–91. |
| [15] | J. P. Legg, African cassava mosaic disease, In: Encyclopedia of virology, Third Edition, Elsevier, 2008, 30–36. |
| [16] |
F. A. Basir, S. Adhurya, M. Banerjee, E. Venturino, S. Ray, Modelling the effect of incubation and latent periods on the dynamics of vector-borne plant viral diseases, Bull. Math. Biol., 82 (2020), 94. https://doi.org/10.1007/s11538-020-00767-2 doi: 10.1007/s11538-020-00767-2
|
| [17] | J. E. Van der Plank, Plant diseases: Epidemics and control, New York: Academic Press, 2013. |
| [18] |
M. J. Jeger, J. Holt, F. van den Bosch, L.V. Madden, Epidemiology of insect-transmitted plant viruses: Modelling disease dynamics and control interventions, Physiol. Entomol., 29 (2004), 291–304. https://doi.org/10.1111/j.0307-6962.2004.00394.x doi: 10.1111/j.0307-6962.2004.00394.x
|
| [19] |
M. Jackson, B. M. Chen-Charpentier, A model of biological control of plant virus propagation with delays, J. Comput. Appl. Math., 330 (2018), 855–865. https://doi.org/10.1016/j.cam.2017.01.005 doi: 10.1016/j.cam.2017.01.005
|
| [20] |
M. Jackson, B. M. Chen-Charpentier, Modeling plant virus propagation with delays, J. Comput. Appl. Math., 309 (2017), 611–621. https://doi.org/10.1016/j.cam.2016.04.024 doi: 10.1016/j.cam.2016.04.024
|
| [21] |
G. Neofytou, Y. N. Kyrychko, K. B. Blyuss, Time-delayed model of immune response in plants, J.Theor. Biol., 389 (2016), 28–39. https://doi.org/10.1016/j.jtbi.2015.10.020 doi: 10.1016/j.jtbi.2015.10.020
|
| [22] |
K. Cooke, P. van den Driessche, X. Zou, Interaction of maturation delay and nonlinear birth in population and epidemic models, J. Math. Biol., 39 (1999), 332–352. https://doi.org/10.1007/s002850050194 doi: 10.1007/s002850050194
|
| [23] |
G. A. Ngwa, On the population dynamics of the malaria vector, Bull. Math. Biol., 68 (2006), 2161–2189. https://doi.org/10.1007/s11538-006-9104-x doi: 10.1007/s11538-006-9104-x
|
| [24] | M. Martcheva, O. Prosper, Unstable dynamics of vector-borne diseases: Modeling through delay-differential equations, In: Dynamic models of infectious diseases, New York: Springer, 2013, 43–75. https://doi.org/10.1007/978-1-4614-3961-5_2 |
| [25] |
M. Banerjee, Y. Takeuchi, Maturation delay for the predators can enhance stable coexistence for a class of prey-predator models, J. Theor. Biol., 412 (2017), 154–171. https://doi.org/10.1016/j.jtbi.2016.10.016 doi: 10.1016/j.jtbi.2016.10.016
|
| [26] |
F. A. Basir, Y. N. Kyrychko, K. B. Blyuss, S. Ray, Effects of vector maturation time on the dynamics of cassava mosaic disease, Bull. Math. Biol., 83 (2021), 87. https://doi.org/10.1007/s11538-021-00921-4 doi: 10.1007/s11538-021-00921-4
|
| [27] |
F. A. Basir, K. B. Blyuss, S. Ray, Modelling the effects of awareness-based interventions to control the mosaic disease of Jatropha curcas, Ecol. Complex, 36 (2018), 92–100. https://doi.org/10.1016/j.ecocom.2018.07.004 doi: 10.1016/j.ecocom.2018.07.004
|
| [28] |
N. Rakshit, F. A. Basir, A. Banerjee, S. Ray, Dynamics of plant mosaic disease propagation and the usefulness of roguing as an alternative biological control, Ecol. Complex., 38 (2019), 15–23. https://doi.org/10.1016/j.ecocom.2019.01.001 doi: 10.1016/j.ecocom.2019.01.001
|
| [29] |
K. B. Blyuss, Control of mosaic disease using microbial biostimulants: Insights from mathematical modelling. Ricerche mat., 69 (2020), 437–455. https://doi.org/10.1007/s11587-020-00508-6 doi: 10.1007/s11587-020-00508-6
|
| [30] |
F. A. Basir, S. Ray, Impact of farming awareness based roguing, insecticide spraying and optimal control on the dynamics of mosaic disease, Ricerche mat., 69, (2020), 393–412. https://doi.org/10.1007/s11587-020-00522-8 doi: 10.1007/s11587-020-00522-8
|
| [31] |
E. Venturino, P. K. Roy, F. A. Basir, A. Datta, A model for the control of the mosaic virus disease in Jatropha curcas plantations, Energ. Ecol. Environ., 1 (2016), 360–369. https://doi.org/10.1007/s40974-016-0033-8 doi: 10.1007/s40974-016-0033-8
|
| [32] |
F. A. Basir, E. Venturino, P. K. Roy, Effects of awareness program for controlling on mosaic disease in Jatropha curcas plantations, Math. Method Appl. Sci., 40 (2017), 2441–2453. https://doi.org/10.1002/mma.4149 doi: 10.1002/mma.4149
|
| [33] |
J. Holt, M. J. Jeger, J. M. Thresh, G. W. Otim-Nape, An epidemiological model incorporating vector population dynamics applied to African cassava mosaic virus disease, J. Appl. Ecol., 34 (1997), 793–806. https://doi.org/10.2307/2404924 doi: 10.2307/2404924
|
| [34] |
S. R. Chant, Studies on the transmission of cassava mosaic virus by Bemisia spp. (Aleyrodidae), Ann. Appl. Biol., 46 (1958), 210–215. https://doi.org/10.1111/j.1744-7348.1958.tb02198.x doi: 10.1111/j.1744-7348.1958.tb02198.x
|
| [35] | H. D. Thurston, Tropical plant diseases, St. Paul: APS press, 1998. |
| [36] | D. Fargette, M. Jeger, C. Fauquet, L. D. Fishpool, Analysis of temporal disease progress of African cassava mosaic Virus, Phytopathology, 84 (1994), 191–198. |
| [37] | C. Fauquet, D. Fargette, African cassava mosaic virus: Etiology, epidemiology, and control, Plant Dis., 74 (1990), 404–411. |
| [38] | J. K. Hale, Theory of functional differential equations, New York: Springer-Verlag, 1977. https://doi.org/10.1007/978-1-4612-9892-2 |
| [39] |
B. Anderson, J. Jackson, M. Sitharam, Descartes' rule of signs revisited, Amer. Math. Mon., 105 (1998), 447–451. https://doi.org/10.2307/3109807 doi: 10.2307/3109807
|
| [40] | S. A. Levin, Descartes rule of signs-how hard can it be? http://sepwww.stanford.edu/oldsep/stew/descartes.pdf |
| [41] |
J. Dushoff, W. Huang, C. Castillo-Chavez, Backwards bifurcations and catastrophe in simple models of fatal diseases, J. Math. Biol., 36 (1998), 227–248. https://doi.org/10.1007/s002850050099 doi: 10.1007/s002850050099
|
| [42] |
C. Castillo-Chavez, B. Song, Dynamical models of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004), 361–404. https://doi.org/10.3934/mbe.2004.1.361 doi: 10.3934/mbe.2004.1.361
|
| [43] |
B. Buonomo, D. Lacitignola, On the backward bifurcation of a vaccination model with nonlinear incidence, Nonlinear Anal.-Model., 16 (2011), 30–46. https://doi.org/10.15388/NA.16.1.14113 doi: 10.15388/NA.16.1.14113
|
| [44] |
A. K. Misra, P. K. Tiwari, P. Chandra, Modeling the control of algal bloom in a lake by applying some external efforts with time delay, Differ. Equ. Dyn. Syst., 29 (2021), 539–568. https://doi.org/10.1007/s12591-017-0383-5 doi: 10.1007/s12591-017-0383-5
|
| [45] |
J. M. Heffernan, R. J. Smith, L. M. Wahl, Perspectives on the basic reproductive ratio, J. R. Soc. Interface, 2 (2005), 281–293. https://doi.org/10.1098/rsif.2005.0042 doi: 10.1098/rsif.2005.0042
|






Figures(7) / Tables(1)

Fahad Al Basir, Konstantin B. Blyuss, Ezio Venturino. Stability and bifurcation analysis of a multi-delay model for mosaic disease transmission[J]. AIMS Mathematics, 2023, 8(10): 24545-24567. doi: 10.3934/math.20231252







 DownLoad:
DownLoad: