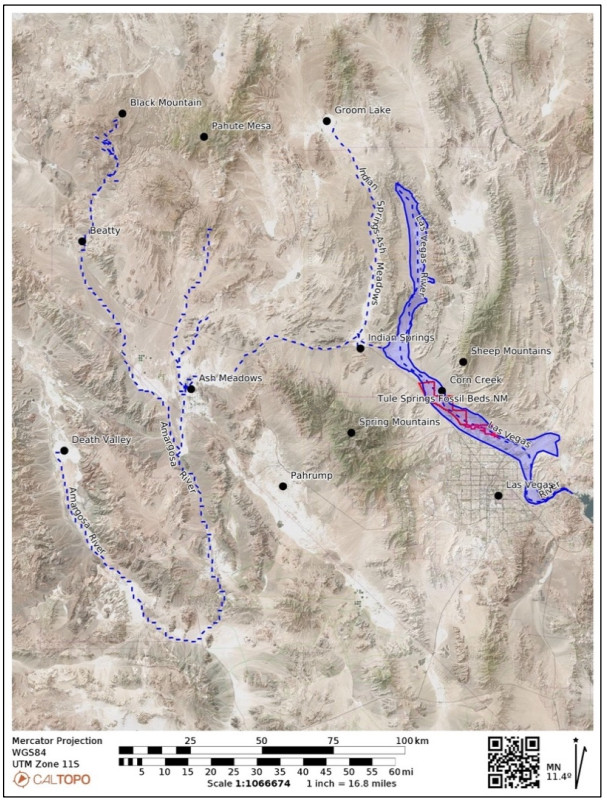
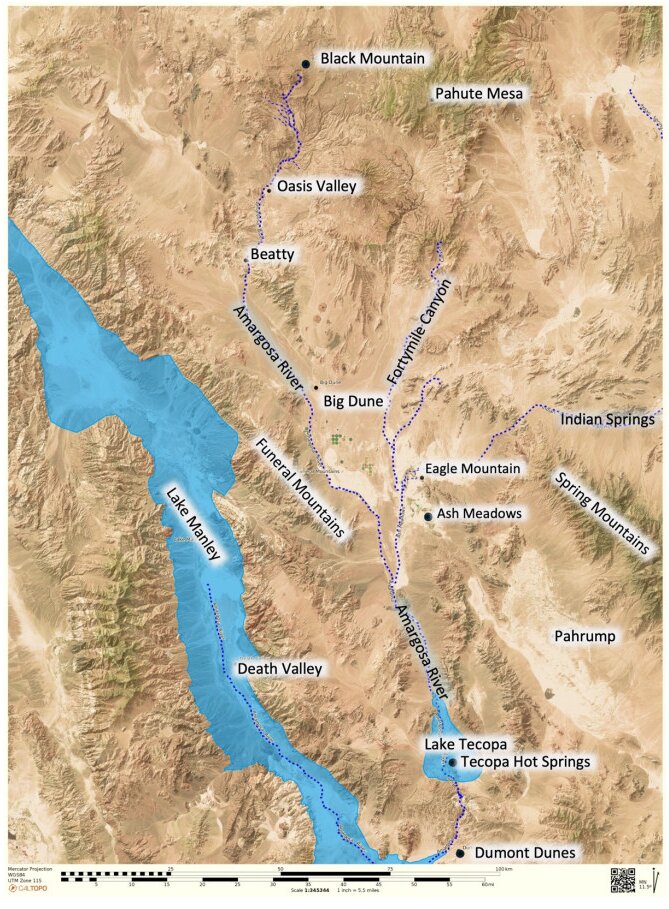
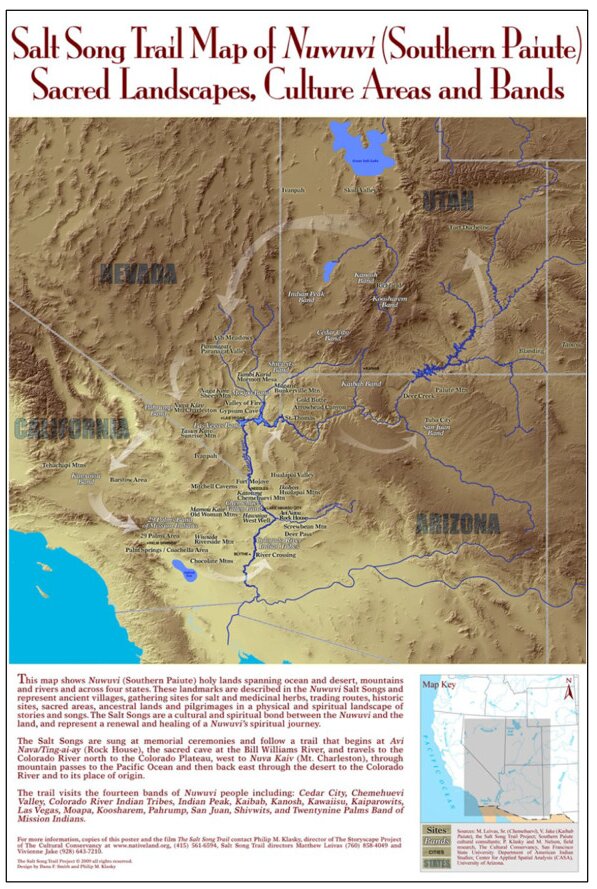
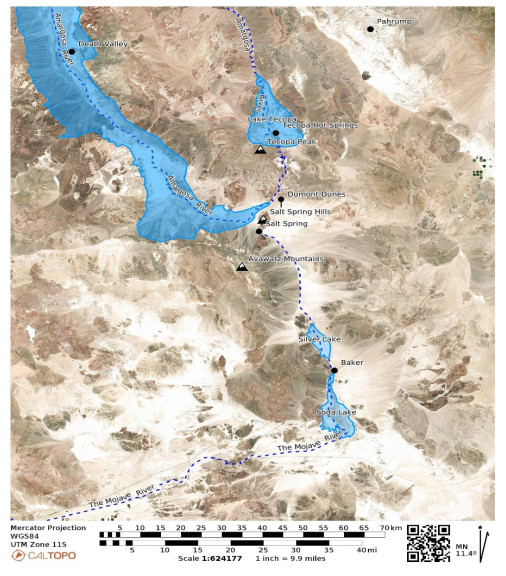
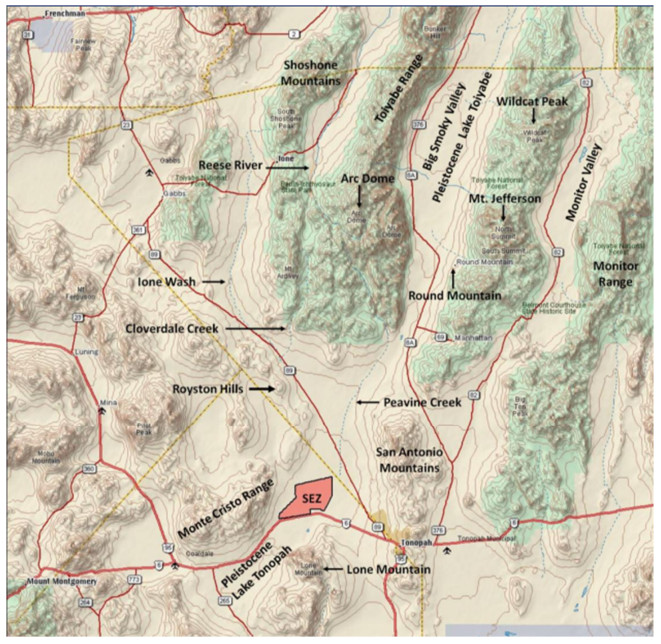
This is a data-based analysis of how Native American interpretations of their distant past are being considered reflecting new science findings. A key science understanding developed over the past 75 years has been that Native people did not occupy North America (or any place in the so-called New World) longer than 12,000 years before present (BP), thus they could neither have experienced nor understood any event in the late Pleistocene interglacial period (128,000 BP to 11,700 BP). As called in this analysis, the Clovis glass ceiling references the popular use of Clovis spear points to represent the earliest signs of humans in North America with dates generally later than 12,000 BP. This analysis engaged with recent science findings that Native people were present in North America up to 40,000 years ago. Opening the science limits of Native presence affords a reinterpretation of the past using extant Native interpretations. As an example, Salt Spring near Death Valley is a component of an ancient Pleistocene heritage landscape that can be reconstructed using geology and Native American interpretations. Native American perspectives were derived from 404 ethnographic interviews with Numic speaking peoples, focused on 24 ancient springs near Death Valley, California, and Las Vegas, Nevada.
Citation: Richard W. Stoffle, Kathleen A. Van Vlack, Heather H. Lim, Alannah Bell, Landon Yarrington. Breaking the Clovis glass ceiling: Native American oral history of the Pleistocene[J]. AIMS Geosciences, 2024, 10(3): 436-459. doi: 10.3934/geosci.2024023
This is a data-based analysis of how Native American interpretations of their distant past are being considered reflecting new science findings. A key science understanding developed over the past 75 years has been that Native people did not occupy North America (or any place in the so-called New World) longer than 12,000 years before present (BP), thus they could neither have experienced nor understood any event in the late Pleistocene interglacial period (128,000 BP to 11,700 BP). As called in this analysis, the Clovis glass ceiling references the popular use of Clovis spear points to represent the earliest signs of humans in North America with dates generally later than 12,000 BP. This analysis engaged with recent science findings that Native people were present in North America up to 40,000 years ago. Opening the science limits of Native presence affords a reinterpretation of the past using extant Native interpretations. As an example, Salt Spring near Death Valley is a component of an ancient Pleistocene heritage landscape that can be reconstructed using geology and Native American interpretations. Native American perspectives were derived from 404 ethnographic interviews with Numic speaking peoples, focused on 24 ancient springs near Death Valley, California, and Las Vegas, Nevada.

| [1] | Stoffle R, Van Vlack K, Johnson H, et al. (2011) Tribally Approved American Indian Ethnographic Analysis of the Proposed Amargosa Valley Solar Energy Zone. Tucson, AZ, USA: Bureau of Applied Research in Anthropology, University of Arizona. Available from: https://solareis.anl.gov/documents/ethnographic/EthnographicAnalysis_AmargosaValley.pdf. |
| [2] |
Bennett M, Bustos D, Pigati J, et al. (2021) Evidence of humans in North America during the Last Glacial Maximum. Science 373:1528–1531. https://doi.org/10.1126/science.abg7586 doi: 10.1126/science.abg7586

|
| [3] |
Pigati JS, Springer KB, Honke JS, et al. (2023) Independent age estimates resolve the controversy of ancient human footprints at White Sands. Science 382:73–75. https://doi.org/10.1126/science.adh5007 doi: 10.1126/science.adh5007
|
| [4] |
Rowe T, Stafford TW, Fisher D, et al. (2022) Human Occupation of the North American Colorado Plateau ∼37,000 Years Ago. Front Ecol Evol 10. https://doi.org/10.3389/fevo.2022.903795 doi: 10.3389/fevo.2022.903795
|
| [5] |
Pitblado BL (2022) On Rehumanizing Pleistocene People of the Western Hemisphere. American Antiquity, 87: 217–235. https://doi.org/10.1017/aaq.2021.120 doi: 10.1017/aaq.2021.120
|
| [6] |
Surovell TA, Allaun SA, Crass BA, et al. (2022) Late date of human arrival to North America: Continental scale differences in stratigraphic integrity of pre-13,000 BP archaeological sites. PLoS ONE 17: e0264092. https://doi.org/10.1371/journal.pone.0264092 doi: 10.1371/journal.pone.0264092
|
| [7] | Deloria V (1997) Red Earth, White Lies: Native Americans and the Myth of Scientific Fact. Golden, CO: Fulcrum Publishing. |
| [8] | Steeves P (2015) Decolonizing Indigenous histories, Pleistocene archaeology sites of the Western Hemisphere. Unpublished Dissertation. |
| [9] | Steeves P (2021) The Indigenous Paleolithic of the Western HemisphereSt. Lincoln, Nebraska: University of Nebraska Press. |
| [10] |
Adams KD, Goebel T, Graf K, et al. (2008) Late Pleistocene and Early Holocene lake-level fluctuations in the Lahontan Basin, Nevada: Implications for the distribution of archaeological sites. Geoarchaeology 23: 608–643. https://doi.org/10.1002/gea.20237 doi: 10.1002/gea.20237
|
| [11] | Malotki E, Wallace H (2011) Columbian mammoth petroglyphs from the San Juan River near Bluff, Utah, United States. Rock Art Res 28: 143–152. https://search.informit.org/doi/10.3316/ielapa.495390102265650 |
| [12] |
Whitley DS (2013) Rock Art Dating and the Peopling of the Americas. J Archaeol 2013: 713159. https://doi.org/10.1155/2013/713159 doi: 10.1155/2013/713159
|
| [13] |
Whitley DS, Dorn RI, Simon JM, et al. (1999) Sally's Rockshelter and the Archaeology of the Vision Quest. Cambridge Archaeol J 9: 221–247. https://doi.org/10.1017/S0959774300015390 doi: 10.1017/S0959774300015390
|
| [14] |
Whitley DS (2024) Ontological Beliefs and Hunter–Gatherer Ritual Landscapes: Native Californian Examples. Religions 15: 123. https://doi.org/10.3390/rel15010123 doi: 10.3390/rel15010123
|
| [15] | Stoffle R, Pickering E, Sittler C, et al. (2017) Ethnographic Overview and Assessment for Canyonlands National Park. Tucson, AZ: University of Arizona. |
| [16] | Stoffle R, Zedeñ o N, Arnold R, et al. (2006) Dá Me Na-Nu-Wu-Tsi: "Our Relations All of Mother Earth" Timber Mountain Ethnographic Report. Tucson, AZ: Bureau of Applied Research in Anthropology, University of Arizona. |
| [17] | Johnson EA, Arlidge SM (2024) Natural Science and Indigenous Knowledge. In: Natural Science and Indigenous Knowledge: The Americas Experience. Cambridge: Cambridge University Press. Available from: https://www.cambridge.org/core/product/DFF680C689A2AE097297BFACEF52F183. |
| [18] |
Whittaker J (1997) Red Earth, White Lies: Native Americans and The Myth of Scientific Fact-Review. Skeptical Inquirer, 21(1). https://doi.org/10.1525/nr.1997.1.1.163 doi: 10.1525/nr.1997.1.1.163
|
| [19] |
Mohawk J (1996) Red Earth, White Lies: Native Americans and the Myth of Scientific Fact. Vine Deloria Jr. American Anthropologist. 98: 650–651. https://doi.org/10.1525/aa.1996.98.3.02a00280 doi: 10.1525/aa.1996.98.3.02a00280
|
| [20] |
Brumble HD (1998) Vine Deloria, Jr., Creationism, and Ethnic Pseudoscience. Am Lit Hist 10: 335–346. https://doi.org/10.1093/alh/10.2.335 doi: 10.1093/alh/10.2.335
|
| [21] | Stoffle R (2007) Cultural Heritage and Resources. In: Fairclough G, Harrison R, Schofield J, editors. The Heritage Reader. London, England: Routledge, 363–372. |
| [22] | Stoffle R (2000) Culture Heritage and Resources. In: Goodman L, editor. Social Impact Analysis: An Applied Anthropology Manual. New York, NY, USA: BERG, 191–232. https://doi.org/10.4324/9781003135838-10 |
| [23] | Sauer C (1925) The Morphology of Landscape. Berkley, CA, USA: University of California, Report No.: 22, 19–53. |
| [24] | Fowler PJ (2003) World Heritage Cultural landscapes. Paris, France: UNESCO World Heritage Centre, 1992–2000. |
| [25] | Stegmeijer E, Veldpaus L (2021) A Research Agenda for Heritage Planning: Perspectives from Europe. Cheltenham, UK: Edward Elgar Publishing. (Elgar Research Agendas). https://doi.org/10.4337/9781788974639.00030 |
| [26] | IUCN (2024) International Union for Conservation of Nature. International Union for Conservation of Nature. Available from: iucn.org. |
| [27] | Mitchell N, Rö ssler M, Tricaud M (2009) World Heritage Cultural Landscapes: A Handbook for Conservation and Management. Paris, France: UNESCO World Heritage Centre. |
| [28] | Stoffle R, Van Vlack K, Johnson H, et al. (2011) Tribally Approved American Indian Ethnographic Analysis of the Proposed Escalante Solar Energy Zone. Tucson, AZ, USA: Bureau of Applied Research in Anthropology, University of Arizona. Available from: https://solareis.anl.gov/documents/ethnographic/EthnographicAnalysis_Millers.pdf. |
| [29] | Stoffle R, Van Vlack K, Lim H (2023) Tiering Southern Paiute Springs and Hydrological Systems. Tucson, AZ, USA: Bureau of Applied Research in Anthropology, University of Arizona. |
| [30] | Stoffle R, Sittler C, Johnson C, et al. (2019) CGTO Writers Committee. Nevada Test and Training Range, Nellis Air Force Base, Legislative Environmental Impact Statement; Native American Ethnographic Study. Washington D.C, USA: Department of Defense. |
| [31] | Stoffle R, Toupal, R, Zedeñ o M (2002) East of Nellis: Cultural Landscapes of the Sheep and Pahranagat Mountain Ranges, An Ethnographic Assessment of American Indian Places and Resources in the Desert National Wildlife Range and the Pahranagat National Wildlife Refuge of Nevada. Tucson, AZ, USA: Bureau of Applied Research in Anthropology, University of Arizona. |
| [32] | Stoffle R, Pittaluga F, Earnest T, et al. (1998) Dewey-Hefley G. Pah Hu Wichi (From Big Spring Running Down): Big Spring Ethnographic Assessment US 95 Corridor Study. Tucson, AZ, USA: Bureau of Applied Research in Anthropology, University of Arizona. |
| [33] | Stoffle, R, Zedeñ o M, Eisenberg A, et al. (2000) Ha'tata (The Backbone of the River): American Indian Ethnographic Studies Regarding the Hoover Dam Bypass Project. Tucson, AZ, USA: Bureau of Applied Research in Anthropology, University of Arizona. |
| [34] | Stoffle R, Van Vlack K, Toupal, R, O'Meara S, Arnold R. American Indians and the Old Spanish Trail. Tucson, AZ: Bureau of Applied Research in Anthropology, University of Arizona; 2008. |
| [35] | Carroll A, Stoffle R, Van Vlack K, et al. (2006) Black Mountain: Traditional Use of a Volcanic Landscape. Prepared for Nellis Air Force Base- Air Command Nevada Test and Training Range Native American Program. Tucson, AZ: University of Arizona. |
| [36] | Stoffle R, Arnold R, Van Vlack K, et al. (2009) Black Mountain: Traditional Uses of a Volcanic Landscape Vol. Ⅱ. Tucson, AZ: Bureau of Applied Research in Anthropology, University of Arizona. |
| [37] | Arnold R, Booth E, Cloquet D, et al. (1999) American Indian Transportation Committee Field Assessment of Cultural Sites Regarding the U.S. Department of Energy: Preapproval Draft Environmental Assessment of Intermodal Transportation of Low-Level Radioactive Waste to the Nevada Test Site. Tucson, AZ: Bureau of Applied Research in Anthropology, University of Arizona. |
| [38] | Stoffle R, Evans M, Halmo D (1989) Native American Plant Resources in the Yucca Mountain Area, Nevada. Ann Arbor, Michigan, USA: Institute for Social Research. https://doi.org/10.2172/137690 |
| [39] | Stoffle R, Chmara-Huff F, Van Vlack K, et al. (2004) Puha Flows from It: The Cultural Landscape Study of the Spring Mountains. Tucson, AZ, USA: Bureau of Applied Research in Anthropology, University of Arizona. |
| [40] |
Anderson DE (2005) Holocene fluvial geomorphology of the Amargosa River through Amargosa Canyon, California. Earth Sci Rev 73: 291–307. https://doi.org/10.1016/j.earscirev.2005.04.010 doi: 10.1016/j.earscirev.2005.04.010
|
| [41] |
Stoffle R, Halmo D, Austin D (1997) Cultural Landscapes and Traditional Cultural Properties: A Southern Paiute View of the Grand Canyon and Colorado River. Am Indian Q 21: 229–249. https://doi.org/10.2307/1185646 doi: 10.2307/1185646
|
| [42] | Cultural Conservancy (2024) Salt Song Trail Project. Salt Song Trail Project. Available from: http://www.nativeland.org/saltsong.html. |
| [43] |
Andreotti B, Bonneau L (2009) Booming Dune Instability. Phys Rev Lett 103: 238001. https://doi.org/10.1103/PhysRevLett.103.238001 doi: 10.1103/PhysRevLett.103.238001
|
| [44] | Haff P (1979) Booming Sands of the Mojave Desert and the Basin and Range Province, California. Pasadena, California: California Institute of Technology. |
| [45] |
Vriend NM, Hunt ML, Clayton RW, et al. (2007) Solving the mystery of booming sand dunes. Geophys Res Lett 34. https://doi.org/10.1029/2007GL030276 doi: 10.1029/2007GL030276
|
| [46] |
Dutta K (2016) Singing Sand Dunes. Resonance 21: 339–351. https://doi.org/10.1007/s12045-016-0333-5 doi: 10.1007/s12045-016-0333-5
|
| [47] |
Enzel Y (1992) Flood Frequency of the Mojave River and the Formation of Late Holocene Playa Lakes, Southern California, USA. Holocene 2: 11–18. https://doi.org/10.1177/095968369200200102 doi: 10.1177/095968369200200102
|
| [48] |
ORE HT, WARREN CN (1971) Late Pleistocene-Early Holocene Geomorphic History of Lake Mojave, California. GSA Bull 82: 2553–2562. https://doi.org/10.1130/0016-7606(1971)82[2553:LPHGHO]2.0.CO;2 doi: 10.1130/0016-7606(1971)82[2553:LPHGHO]2.0.CO;2
|
| [49] |
Enzel Y, Brown WJ, Anderson RY, et al. (1992) Short-Duration Holocene Lakes in the Mojave River Drainage Basin, Southern California. Quat Res 38: 60–73. https://doi.org/10.1016/0033-5894(92)90030-M doi: 10.1016/0033-5894(92)90030-M
|
| [50] | Earle D (2005) The Mojave River and the Central Mojave Desert: Native Settlement, Travel, and Exchange in the Eighteenth and Nineteenth Centuries. J Calif Great Basin Anthropol 25: 1–38. |
| [51] | Bean LJ (1978) Social Organization. In: Sturtevant W, editor. California, Handbook of North American Indians. Washington D.C, USA: Smithsonian Institution Press, 673–682. |
| [52] | Kroeber A (1925) Handbook of the Indians of California. Washington D.C, USA: Bureau of American Ethnology, Vol. 78. |
| [53] |
Gould D, Glowacka M (2004) Nagotooh(Gahni): The Bonding between Mother and Child in Shoshoni Tradition. Ethnology 43: 185–191. https://doi.org/10.2307/3773953 doi: 10.2307/3773953
|
| [54] | Solar PEIS (2012) Ethnographic Analyses for Nine Proposed SEZs. Social Energy Development Programmatic EIS Information Center[Internet]. Ethnographic Analyses for Nine Proposed SEZs. Social Energy Development Programmatic EIS Information Center. Available from: https://solareis.anl.gov/documents/ethnographic/index.cfm. |
| [55] |
Mughal MAZ (2023) The western and non-western dichotomization of time in anthropology. Int J Anthropol Ethnol 7: 7. https://doi.org/10.1186/s41257-023-00086-z doi: 10.1186/s41257-023-00086-z
|
| [56] | Mughal MAZ (2014) Time, Space and Social Change in Rural Pakistan: An Ethnographic Study of Jhokwala Village, Lodhran District. Dissertation. Durham, England: Durham University. |
| [57] | Jennings F (1976) The Invasion of America: Indians, Colonialism, and the Cant of Conquest. Chapel Hill, NC: University of North Carolina Press. |
| [58] | Stoffle R (2022) Living Stone Bridges: Epistemological Divides in Heritage Environmental Communication. In: Sjö lander-Lindqvist A, Murin I, Dove M, editors. Anthropological Perspectives on Environmental Communication. Switzerland: Palgrave Macmillan, 149–73. Available from: https://link.springer.com/book/10.1007/978-3-030-78040-1. |
| [59] | Johnson C (2023) Ancient Footprints Upend Timeline of Humans' Arrival in North America.". Washington Post. Available from: https://www.washingtonpost.com/science/2023/10/05/oldest-human-footprint-americas-white-sands/. |




Figures(9) / Tables(1)

Richard W. Stoffle, Kathleen A. Van Vlack, Heather H. Lim, Alannah Bell, Landon Yarrington. Breaking the Clovis glass ceiling: Native American oral history of the Pleistocene[J]. AIMS Geosciences, 2024, 10(3): 436-459. doi: 10.3934/geosci.2024023







 DownLoad:
DownLoad: