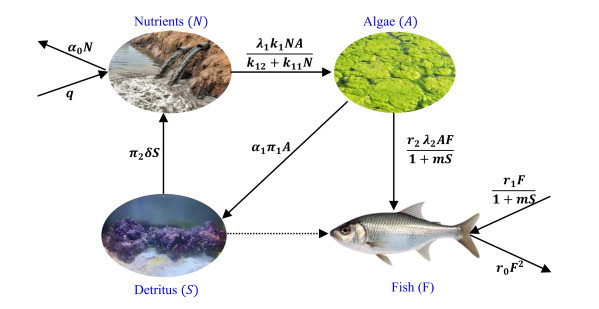
Algal blooms pose a significant threat to the ecological integrity and biodiversity in aquatic ecosystems. In lakes, enriched with nutrients, these blooms result in overgrowth of periphyton, leading to biological clogging, oxygen depletion, and ultimately a decline in ecosystem's health and water quality. In this article, we presented a mathematical model centered around the role of aquatic species (specifically fish population) to alleviate algal blooms. The model analysis revealed significant shifts in dynamics, shedding light on the effectiveness of fish-mediated sustainability strategies to control algal proliferation. Notably, our study identified critical thresholds and regime transitions through the observation of saddle-node bifurcation within the proposed mathematical model. To validate our analytical findings, we have conducted numerical simulations, which provided robust evidence for the resilience of the ecosystem under different scenarios.
Citation: Mohammad Sajid, Arvind Kumar Misra, Ahmed S. Almohaimeed. Modeling the role of fish population in mitigating algal bloom[J]. Electronic Research Archive, 2024, 32(10): 5819-5845. doi: 10.3934/era.2024269
Algal blooms pose a significant threat to the ecological integrity and biodiversity in aquatic ecosystems. In lakes, enriched with nutrients, these blooms result in overgrowth of periphyton, leading to biological clogging, oxygen depletion, and ultimately a decline in ecosystem's health and water quality. In this article, we presented a mathematical model centered around the role of aquatic species (specifically fish population) to alleviate algal blooms. The model analysis revealed significant shifts in dynamics, shedding light on the effectiveness of fish-mediated sustainability strategies to control algal proliferation. Notably, our study identified critical thresholds and regime transitions through the observation of saddle-node bifurcation within the proposed mathematical model. To validate our analytical findings, we have conducted numerical simulations, which provided robust evidence for the resilience of the ecosystem under different scenarios.

| [1] |
B. Bhagowati K. U. Ahamad, A review on lake eutrophication dynamics and recent developments in lake modeling, Ecohydrol. Hydrobiol., 19 (2019), 155–166. https://doi.org/10.1016/j.ecohyd.2018.03.002 doi: 10.1016/j.ecohyd.2018.03.002

|
| [2] |
H. Wang, H. Wang, Mitigation of lake eutrophication: Loosen nitrogen control and focus on phosphorus abatement, Prog. Nat. Sci., 19 (2009), 1445–1451. https://doi.org/10.1016/j.pnsc.2009.03.009 doi: 10.1016/j.pnsc.2009.03.009
|
| [3] |
A. K. Misra, P. Chandra, V. Raghavendra, Modeling the depletion of dissolved oxygen in a lake due to algal bloom: Effect of time delay, Adv. Water Resour., 34 (2011), 1232–1238. https://doi.org/10.1016/j.advwatres.2011.05.010 doi: 10.1016/j.advwatres.2011.05.010
|
| [4] |
X. Li, T. Yan, R. Yu, M. Zhou, A review of Karenia mikimotoi: Bloom events, physiology, toxicity and toxic mechanism, Harmful Algae, 90 (2019), 101702. https://doi.org/10.1016/j.hal.2019.101702 doi: 10.1016/j.hal.2019.101702
|
| [5] |
M. Pal, P. J. Yesankar, A. Dwivedi, A. Qureshi, Biotic control of harmful algal blooms (HABs): A brief review, J. Environ. Manage., 268 (2020), 110687. https://doi.org/10.1016/j.jenvman.2020.110687 doi: 10.1016/j.jenvman.2020.110687
|
| [6] |
L. M. Grattan, S. Holobaugh, J. G. Morris Jr, Harmful algal blooms and public health, Harmful Algae, 57 (2016), 2–8. https://doi.org/10.1016/j.hal.2016.05.003 doi: 10.1016/j.hal.2016.05.003
|
| [7] | S. Xiang, Y. Han, C. Jiang, M. Li, L. Wei, J. Fu, et al., Composite biologically active filter (BAF) with zeolite, granular activated carbon, and suspended biological carrier for treating algae-laden raw water, J. Water Process Eng., 42, (2021), 102188. https://doi.org/10.1016/j.jwpe.2021.102188 |
| [8] |
Y. Peng, W. Zhang, X. Yang, Z. Zhang, G. Zhu, S. Zhou, Current status and prospects of algal bloom early warning technologies: A review, J. Environ. Manage., 349 (2024), 119510. https://doi.org/10.1016/j.jenvman.2023.119510 doi: 10.1016/j.jenvman.2023.119510
|
| [9] |
Y. Peng, X. Xiao, B. Ren, Z. Zhang, J. Luo, X. Yang, et al., Biological activity and molecular mechanism of inactivation of Microcystis aeruginosa by ultrasound irradiation, J. Hazard. Mater., 468 (2024), 133742. https://doi.org/10.1016/j.jhazmat.2024.133742 doi: 10.1016/j.jhazmat.2024.133742
|
| [10] |
C. Chen, X. Pang, Y. Wang, M. Kong, L. Long, M. Xu, et al., Antioxidant responses and microcystins accumulation in Corbicula fluminea following the control of algal blooms using chitosan-modified clays, J. Soils Sediments, 21 (2021), 3505–3514. https://doi.org/10.1007/s11368-021-03022-w doi: 10.1007/s11368-021-03022-w
|
| [11] |
K. Liu, L. Jiang, J. Yang, S. Ma, K. Chen, Y. Zhang, et al., Comparison of three flocculants for heavy cyanobacterial bloom mitigation and subsequent environmental impact, J. Oceanol. Limnol., 40 (2022), 1764–1773. https://doi.org/10.1007/s00343-022-1351-7 doi: 10.1007/s00343-022-1351-7
|
| [12] |
D. E. Berthold, A. Elazar, F. Lefler, C. Marble, H. D. Laughinghouse IV, Control of algal growth on greenhouse surfaces using commercial algaecides, Sci. Agri., 78 (2020), e20180292. https://doi.org/10.1590/1678-992x-2018-0292 doi: 10.1590/1678-992x-2018-0292
|
| [13] |
P. Laue, H. Bahrs, S. Chakrabarti, C. E. Steinberg, Natural xenobiotics to prevent cyanobacterial and algal growth in freshwater: Contrasting efficacy of tannic acid, gallic acid, and gramine, Chemosphere, 104 (2014), 212–220. https://doi.org/10.1016/j.chemosphere.2013.11.029 doi: 10.1016/j.chemosphere.2013.11.029
|
| [14] |
R. Sun, P. Sun, J. Zhang, S. Esquivel-Elizondo, Y. Wu, Microorganisms-based methods for harmful algal blooms control: A review, Bioresour. Technol., 248 (2018), 12–20. https://doi.org/10.1016/j.biortech.2017.07.175 doi: 10.1016/j.biortech.2017.07.175
|
| [15] | I. Domaizon, J. Devaux, A new approach in biomanipulation techniques: Use of a phytoplanktivorous fish, the silver carp (Hypophthalmichthys molitrix), Anne Biol., 38 (1999). https://doi.org/10.1016/S0003-5017(99)80028-2 |
| [16] |
M. K. Ekvall, P. Urrutia-Cordero, L. A. Hansson, Linking cascading effects of fish predation and zooplankton grazing to reduced cyanobacterial biomass and toxin levels following biomanipulation, PloS One, 9 (2014), 112956. https://doi.org/10.1371/journal.pone.0112956 doi: 10.1371/journal.pone.0112956
|
| [17] |
G. W. Waajen, N. C. Van Bruggen, L. M. D. Pires, W. Lengkeek, M. Lürling, Biomanipulation with quagga mussels (Dreissena rostriformis bugensis) to control harmful algal blooms in eutrophic urban ponds, Ecol. Eng., 90 (2016), 141–150. https://doi.org/10.1016/j.ecoleng.2016.01.036 doi: 10.1016/j.ecoleng.2016.01.036
|
| [18] | A. A. Voinov, A. P. Tonkikh, Qualitative model of eutrophication in macrophyte lakes, Ecol. Model., 35, (1987), 211–226. https://doi.org/10.1016/0304-3800(87)90113-X |
| [19] | M. Scheffer, S. Carpenter, J. A. Foley, C. Folke, B. Walker, Catastrophic shifts in ecosystems, Nature, 413 (2001) 591–596. https://doi.org/10.1038/35098000 |
| [20] |
X. Mao, X. Wei, D. Yuan, Y. Jin, X. Jin, An ecological-network-analysis based perspective on the biological control of algal blooms in Ulansuhai Lake, China, Ecol. Model., 386 (2018), 11–19. https://doi.org/10.1016/j.ecolmodel.2018.07.020 doi: 10.1016/j.ecolmodel.2018.07.020
|
| [21] |
A. K. Misra, Mathematical modeling and analysis of eutrophication of water bodies caused by nutrients, Nonlinear Anal. Model. Control, 12 (2007), 511–524. https://doi.org/10.15388/NA.2007.12.4.14683 doi: 10.15388/NA.2007.12.4.14683
|
| [22] |
P. K. Tiwari, S. Samanta, F. Bona, E. Venturino, A. K. Misra, The time delays influence on the dynamical complexity of algal blooms in the presence of bacteria, Ecol. Complex., 39 (2019), 100769. https://doi.org/10.1016/j.ecocom.2019.100769 doi: 10.1016/j.ecocom.2019.100769
|
| [23] | J. B. Shukla, A. K. Misra, P. Chandra, Mathematical modeling and analysis of the depletion of dissolved oxygen in eutrophied water bodies affected by organic pollutants, Nonlinear Anal. Real World Appl., 9, (2008), 1851–1865. https://doi.org/10.1016/j.nonrwa.2007.05.016 |
| [24] |
H. Yan, D. Wu, Y. Huang, G. Wang, M. Shang, J. Xu, et al., Water eutrophication assessment based on rough set and multidimensional cloud model, Chemometr Intell. Lab. Syst., 164 (2017), 103–112. https://doi.org/10.1016/j.chemolab.2017.02.005 doi: 10.1016/j.chemolab.2017.02.005
|
| [25] |
J. Zhang, S. E. Jorgensen, M. Beklioglu, O. Ince, Hysteresis in vegetation shift–Lake Mogan prognoses, Ecol. Model., 164 (2003), 227–238. https://doi.org/10.1016/S0304-3800(03)00050-4 doi: 10.1016/S0304-3800(03)00050-4
|
| [26] | C. Castillo-Garsow, G. Jordan-Salivia, A. Rodriguez-Herrera, Mathematical models for the dynamics of tobacco use, recovery and relapse, Biometrics Unit Technical Reports, Number BU-1505-M, (1997). |
| [27] |
Q. An, H. Wang, X. Wang, Fish survival subject to algal bloom: Resource-based growth models with algal digestion delay and detritus-nutrient recycling delay, Ecol. Modell., 491 (2024), 110672. https://doi.org/10.1016/j.ecolmodel.2024.110672 doi: 10.1016/j.ecolmodel.2024.110672
|
| [28] |
W. J. O'Brien, The dynamics of nutrient limitation of phytoplankton algae: A model reconsidered, Ecology, 55 (1974), 135–141. https://doi.org/10.2307/1934626 doi: 10.2307/1934626
|
| [29] |
A. Huppert, B. Blasius, R. Olinky, L. Stone, A model for seasonal phytoplankton blooms, J. Theor. Biol., 236 (2005), 276–290. https://doi.org/10.1016/j.jtbi.2005.03.012 doi: 10.1016/j.jtbi.2005.03.012
|
| [30] |
M. Chen, M. Fan, R. Liu, X. Wang, X. Yuan, H. Zhu, The dynamics of temperature and light on the growth of phytoplankton, J. Theor. Biol., 385 (2015), 8–19. https://doi.org/10.1016/j.jtbi.2015.07.039 doi: 10.1016/j.jtbi.2015.07.039
|
| [31] |
S. Zhao, S. Yuan, H. Wang, Threshold behavior in a stochastic algal growth model with stoichiometric constraints and seasonal variation, J. Differ. Equations, 268 (2020), 5113–5139. https://doi.org/10.1016/j.jde.2019.11.004 doi: 10.1016/j.jde.2019.11.004
|









Figures(10) / Tables(1)
Mohammad Sajid, Arvind Kumar Misra, Ahmed S. Almohaimeed. Modeling the role of fish population in mitigating algal bloom[J]. Electronic Research Archive, 2024, 32(10): 5819-5845. doi: 10.3934/era.2024269







 DownLoad:
DownLoad: