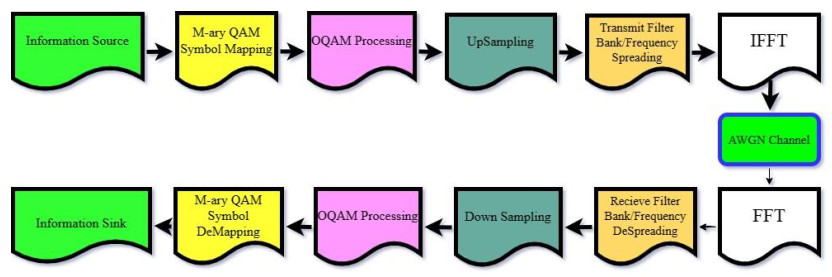
Offset quadrature amplitude modulation–based filter bank multicarrier (FBMC-OQAM) method is a promising technology for future wireless communication systems. It offers several advantages over traditional orthogonal frequency-division multiplexing (OFDM) modulation, including higher spectral efficiency, lower out-of-band emission, and improved robustness to time-frequency selective channels. Polar codes, a new class of error-correcting codes, have received much attention recently due to their ability to achieve the Shannon limit with practical decoding complexity. This paper analyzed and investigated the error rate performance of polar-coded FBMC-OQAM systems. Our results show that applying polar codes to FBMC-OQAM systems significantly improves the error rate. In addition, we found that employing random code interleavers can yield additional coding gains of up to 0.75 dB in additive white Gaussian noise (AWGN) and 2 dB in Nakagami-m fading channels. Our findings suggest that polar-coded FBMC-OQAM is a promising combination for future wireless communication systems. We also compared turbo-coded FBMC-OQAM for short code lengths, and our simulations showed that polar codes exhibit comparable error-correcting capabilities. These results will be of interest to researchers and engineers working on the advancement of future wireless communication systems.
Citation: Tadele A. Abose, Fanuel O. Ayana, Thomas O. Olwal, Yihenew W. Marye. BER performance analysis of polar-coded FBMC/OQAM in the presence of AWGN and Nakagami-m fading channel[J]. AIMS Electronics and Electrical Engineering, 2024, 8(3): 311-331. doi: 10.3934/electreng.2024014
Offset quadrature amplitude modulation–based filter bank multicarrier (FBMC-OQAM) method is a promising technology for future wireless communication systems. It offers several advantages over traditional orthogonal frequency-division multiplexing (OFDM) modulation, including higher spectral efficiency, lower out-of-band emission, and improved robustness to time-frequency selective channels. Polar codes, a new class of error-correcting codes, have received much attention recently due to their ability to achieve the Shannon limit with practical decoding complexity. This paper analyzed and investigated the error rate performance of polar-coded FBMC-OQAM systems. Our results show that applying polar codes to FBMC-OQAM systems significantly improves the error rate. In addition, we found that employing random code interleavers can yield additional coding gains of up to 0.75 dB in additive white Gaussian noise (AWGN) and 2 dB in Nakagami-m fading channels. Our findings suggest that polar-coded FBMC-OQAM is a promising combination for future wireless communication systems. We also compared turbo-coded FBMC-OQAM for short code lengths, and our simulations showed that polar codes exhibit comparable error-correcting capabilities. These results will be of interest to researchers and engineers working on the advancement of future wireless communication systems.

| [1] | Bizaki HK (2016) Towards 5G wireless networks: a physical layer perspective. BoD–Books on Demand. https://doi.org/10.5772/63098 |
| [2] | Demir AF, Elkourdi M, Ibrahim M, Arslan H (2019) Waveform design for 5G and beyond. arXiv preprint arXiv: 1902.05999. https://doi.org/10.1002/9781119333142.ch2 |
| [3] |
Chang RW (1966) High-speed multichannel data transmission with bandlimited orthogonal signals. Bell Syst Tech J 45: 1775‒1796. https://doi.org/10.1002/j.1538-7305.1966.tb02435.x doi: 10.1002/j.1538-7305.1966.tb02435.x

|
| [4] |
Saltzberg B (1967) Performance of an efficient parallel data transmission system. IEEE Transactions on Communication Technology 15: 805‒811. https://doi.org/10.1109/TCOM.1967.1089674 doi: 10.1109/TCOM.1967.1089674
|
| [5] | Jiang T, Chen D, Ni C, Qu D (2017) OQAM/FBMC for future wireless communications: Principles, technologies and applications. Academic Press. https://doi.org/10.1016/B978-0-12-813557-0.00010-3 |
| [6] |
Nissel R, Rupp M (2018) Pruned DFT-spread FBMC: Low PAPR, low latency, high spectral efficiency. IEEE T Commun 66: 4811‒4825. https://doi.org/10.1109/TCOMM.2018.2837130 doi: 10.1109/TCOMM.2018.2837130
|
| [7] |
Wang Y, Guo Q, Xiang J, Wang L, Liu Y (2024) Bi-orthogonality recovery and MIMO transmission for FBMC Systems based on non-sinusoidal orthogonal transformation. Signal Processing 109427. https://doi.org/10.1016/j.sigpro.2024.109427 doi: 10.1016/j.sigpro.2024.109427
|
| [8] |
Arikan E (2009) Channel polarization: A method for constructing capacity-achieving codes for symmetric binary-input memoryless channels. IEEE T Inform Theory 55: 3051‒3073. https://doi.org/10.1109/TIT.2009.2021379 doi: 10.1109/TIT.2009.2021379
|
| [9] |
Ali MH, Al-Rubaye GA (2024) Performance Evaluation of 5G New Radio Polar Code over Different Multipath Fading Channel Models. International Journal of Intelligent Engineering & Systems 17. https://doi.org/10.22266/ijies2024.0430.36 doi: 10.22266/ijies2024.0430.36
|
| [10] |
Meenalakshmi M, Chaturvedi S, Dwivedi VK (2024) Enhancing channel estimation accuracy in polar-coded MIMO–OFDM systems via CNN with 5G channel models. AEU-Int J Electron Commun 173: 155016. https://doi.org/10.1016/j.aeue.2023.155016 doi: 10.1016/j.aeue.2023.155016
|
| [11] |
Vangala H, Hong Y, Viterbo E (2015) Efficient algorithms for systematic polar encoding. IEEE Commun Lett 20: 17‒20. https://doi.org/10.1109/LCOMM.2015.2497220 doi: 10.1109/LCOMM.2015.2497220
|
| [12] |
Hall EK, Wilson SG (1998) Design and analysis of turbo codes on Rayleigh fading channels. IEEE J Sel Areas Commun 16: 160‒174. https://doi.org/10.1109/49.661105 doi: 10.1109/49.661105
|
| [13] |
Liu L, Ling C (2016) Polar codes and polar lattices for independent fading channels. IEEE T Commun 64: 4923‒4935. https://doi.org/10.1109/TCOMM.2016.2613109 doi: 10.1109/TCOMM.2016.2613109
|
| [14] |
Si H, Koyluoglu OO, Vishwanath S (2014) Polar coding for fading channels: Binary and exponential channel cases. IEEE T Commun 62: 2638‒2650. https://doi.org/10.1109/TCOMM.2014.2345399 doi: 10.1109/TCOMM.2014.2345399
|
| [15] | Trifonov P (2015) Design of polar codes for Rayleigh fading channel. 2015 international symposium on wireless communication systems (ISWCS) 331‒335. IEEE. https://doi.org/10.1109/ISWCS.2015.7454357 |
| [16] | Rafik Z, Hmaied S, Daniel R, Yahia M (2017) BER analysis of FBMC-OQAM systems with Phase Estimation Error. IET Communications. |
| [17] |
Marina P, Isnawati AF, Afandi MA (2020) Performance analysis of FBMC O-QAM system using varied modulation level. Jurnal Infotel 12: 45‒51. https://doi.org/10.20895/infotel.v12i2.482 doi: 10.20895/infotel.v12i2.482
|
| [18] | BS R (2021) Performance Analysis of OFDM, FBMC and UFMC Modulation Schemes for 5G Mobile Communication MIMO systems. Proceedings of the International Conference on IoT Based Control Networks & Intelligent Systems-ICICNIS. |
| [19] |
Hassan ES (2024) Performance enhancement and PAPR reduction for MIMO based QAM-FBMC systems. Plos one 19: e0296999. https://doi.org/10.1371/journal.pone.0296999 doi: 10.1371/journal.pone.0296999
|
| [20] |
Al-Amaireh H, Kollár Z (2022) Low complexity PPN-FBMC receivers with improved sliding window equalizers. Phys Commun 54: 101795. https://doi.org/10.1016/j.phycom.2022.101795 doi: 10.1016/j.phycom.2022.101795
|
| [21] |
Wang Y, Guo Q, Xiang J, Liu Y (2023) Doubly selective channel estimation and equalization based on ICI/ISI mitigation for OQAM-FBMC systems. Phys Commun 59: 102120. https://doi.org/10.1016/j.phycom.2023.102120 doi: 10.1016/j.phycom.2023.102120
|
| [22] | Malkamaki E (1992) Binary and multilevel offset QAM, spectrum efficient modulation schemes for personal communications. In [1992 Proceedings] Vehicular Technology Society 42nd VTS Conference-Frontiers of Technology 325‒328. IEEE. |
| [23] | Bellanger M, Le Ruyet D, Roviras D, Terré M, Nossek J, Baltar L, et al. (2010) FBMC physical layer: a primer. PHYDYAS, January 25: 7‒10. |
| [24] |
Choi JM, Oh Y, Lee H, Seo JS (2017) Pilot-aided channel estimation utilizing intrinsic interference for FBMC/OQAM systems. IEEE T Broadcast 63: 644‒655. https://doi.org/10.1109/TBC.2017.2711143 doi: 10.1109/TBC.2017.2711143
|
| [25] |
Yu B, Hu S, Sun P, Chai S, Qian C, Sun C (2016) Channel estimation using dual-dependent pilots in FBMC/OQAM systems. IEEE Commun Lett 20: 2157‒2160. https://doi.org/10.1109/LCOMM.2016.2599882 doi: 10.1109/LCOMM.2016.2599882
|
| [26] | Bedoui A, Et-tolba M (2020) A neuro-fuzzy based detection approach for HARQ-CC in FBMC-OQAM systems. 2020 9th IFIP international conference on performance evaluation and modeling in wireless networks (PEMWN) 1‒7. IEEE. https://doi.org/10.23919/PEMWN50727.2020.9293073 |
| [27] |
Doré JB, Gerzaguet R, Cassiau N, Ktenas D (2017) Waveform contenders for 5G: Description, analysis and comparison. Phys Commun 24: 46‒61. https://doi.org/10.1016/j.phycom.2017.05.004 doi: 10.1016/j.phycom.2017.05.004
|
| [28] |
Abose TA, Olwal TO, Mohammed MM, Hassen MR (2024) Performance analysis of insertion loss incorporated hybrid precoding for massive MIMO. AIMS Electronics and Electrical Engineering 8: 187‒210. https://doi.org/10.3934/electreng.2024008 doi: 10.3934/electreng.2024008
|
| [29] | Abose TA, Mariye YW, Demissie AM, Olwal TO (2023) Energy Efficiency Enhancement of Ultra Dense Multiuser MIMO System by Using Wyner Model. 2023 14th International Conference on Computing Communication and Networking Technologies (ICCCNT) 1‒6. IEEE. https://doi.org/10.1109/ICCCNT56998.2023.10308201 |
| [30] | TS 38.212 NR; Multiplexing and channel coding, 3GPP, 2018. |
| [31] |
Balatsoukas-Stimming A, Parizi MB, Burg A (2015) LLR-based successive cancellation list decoding of polar codes. IEEE T Signal Process 63: 5165‒5179. https://doi.org/10.1109/TSP.2015.2439211 doi: 10.1109/TSP.2015.2439211
|
| [32] | Afisiadis O, Balatsoukas-Stimming A, Burg A (2014) A low-complexity improved successive cancellation decoder for polar codes. 2014 48th Asilomar Conference on Signals, Systems and Computers 2116‒2120. IEEE. https://doi.org/10.1109/ACSSC.2014.7094848 |
| [33] | Arıkan E (2010) Polar codes: A pipelined implementation. Proc 4th ISBC 2010: 11‒14. |
| [34] | Kahraman S, Ç elebi ME (2012) Code based efficient maximum-likelihood decoding of short polar codes. 2012 IEEE International Symposium on Information Theory Proceedings 1967‒1971. IEEE. https://doi.org/10.1109/ISIT.2012.6283643 |
| [35] | Berrou C, Glavieux A, Thitimajshima P (1993) Near Shannon limit error-correcting coding and decoding: Turbo-codes. 1. Proceedings of ICC'93-IEEE International Conference on Communications 2: 1064‒1070. IEEE. |
| [36] |
Benedetto S, Divsalar D, Montorsi G, Pollara F (1996) A soft-input soft-output maximum a posteriori (MAP) module to decode parallel and serial concatenated codes. TDA progress report 42: 1‒20. https://doi.org/10.1109/4234.552145 doi: 10.1109/4234.552145
|
| [37] |
Viterbi AJ (1998) An intuitive justification and a simplified implementation of the MAP decoder for convolutional codes. IEEE J Sel Areas Commun 16: 260‒264. https://doi.org/10.1109/49.661114 doi: 10.1109/49.661114
|
| [38] | Nakagami M (1960) The m-distribution—A general formula of intensity distribution of rapid fading. Statistical methods in radio wave propagation 3‒36. Pergamon. https://doi.org/10.1016/B978-0-08-009306-2.50005-4 |
| [39] | Abose TA, Megersa KT, Jember KA, Kejela DC, Daka ST, Dinagde MB (2023) Performance Comparison of M-ary Phase Shift Keying and M-ary Quadrature Amplitude Modulation Techniques Under Fading Channels. International Conference on Communication and Intelligent Systems 235‒245. Singapore: Springer Nature Singapore. https://doi.org/10.1007/978-981-97-2079-8_19 |








Figures(9) / Tables(6)

Tadele A. Abose, Fanuel O. Ayana, Thomas O. Olwal, Yihenew W. Marye. BER performance analysis of polar-coded FBMC/OQAM in the presence of AWGN and Nakagami-m fading channel[J]. AIMS Electronics and Electrical Engineering, 2024, 8(3): 311-331. doi: 10.3934/electreng.2024014







 DownLoad:
DownLoad: