Citation: Arianna Bozzolo, Diego Pizzeghello, Alessandra Cardinali, Ornella Francioso, Serenella Nardi. Effects of moderate and high rates of biochar and compost on grapevine growth in a greenhouse experiment[J]. AIMS Agriculture and Food, 2017, 2(1): 113-128. doi: 10.3934/agrfood.2017.1.113

| [1] |
Ramos MC (2016) Soil losses in rainfed Mediterranean vineyards under climate change scenarios. The effects of drainage terraces. AIMS Agric Food 1: 124-143. doi: 10.3934/agrfood.2016.2.124

|
| [2] |
Pérez FL (2016) Viticultural practices in Jumilla (Murcia, Spain): a case study of agriculture and adaptation to natural landscape processes in a variable and changing climate. AIMS Agric Food 1: 265-293. doi: 10.3934/agrfood.2016.3.265
|
| [3] | Reynolds AG (2010) Managing Wine Quality: Viticulture and Wine Quality. Woodhead Publishing. |
| [4] | Tesic D, Keller M, Hutton RJ (2007) Influence of vineyard floor management practices on grapevine vegetative growth, yield, and fruit composition. Am J Enol Vitic 58: 1-11. |
| [5] |
Salomé C, Coll P, Lardo E, et al. (2016) The soil quality concept as a framework to assess management practices in vulnerable agroecosystems: A case study in Mediterranean vineyards. Ecol Ind 61: 456-465. doi: 10.1016/j.ecolind.2015.09.047
|
| [6] |
Menichetti L, Ekblad A, Kätterer T (2015) Contribution of roots and amendments to soil carbon accumulation within the soil profile in a long-term field experiment in Sweden. Agr Ecosyst Environ 200: 79-87. doi: 10.1016/j.agee.2014.11.003
|
| [7] |
Vaccaro S, Muscolo A, Pizzeghello D, et al. (2009) Effect of a compost and its water-soluble fractions on key enzymes of nitrogen metabolism in maize seedlings. J Agric Food Chem 57: 11267-11276. doi: 10.1021/jf901808s
|
| [8] |
Vaughn SF, Deppe NA, Palmquist DE, et al. (2011) Extracted sweet corn tassels as a renewable alternative to peat in greenhouse substrates. Ind Crops Prod 33: 514-517. doi: 10.1016/j.indcrop.2010.10.034
|
| [9] | Cao Y, Ma Y, Guo D, et al. (2017) Chemical properties and microbial responses to biochar and compost amendments in the soil under continuous watermelon cropping. Plant Soil Environ 63: 1-7 |
| [10] |
Cardinali A, Pizzeghello D, Zanin G (2015) Fatty acid methyl ester (FAME) succession in different substrates as affected by the co-application of three pesticides. Plos One 10: e0145501. doi: 10.1371/journal.pone.0145501
|
| [11] |
Steiner C, Harttung T (2014) Biochar as growing media additive and peat substitute. Solid Earth 5: 995-999. doi: 10.5194/se-5-995-2014
|
| [12] |
Spokas KA, Cantrell KB, Novak JM, et al. (2012) Biochar: a synthesis of its agronomic impact beyond carbon sequestration. J Environ Qual 41: 973-989. doi: 10.2134/jeq2011.0069
|
| [13] | Laird DA (2008) The charcoal vision: a win-win-win scenario for simultaneously producing bioenergy, permanently sequestration carbon, while improving soil and water quality. Agron J 100: 178-181. |
| [14] |
Van Zwieten L, Kimber S, Morris S, et al. (2010) Effects of biochar from slow pyrolysis of papermill waste on agronomic performance and soil fertility. Plant Soil 327: 235-246. doi: 10.1007/s11104-009-0050-x
|
| [15] | Laird DA, Rogovska NP, Garcia-Perez M, et al. (2011) Pyrolysis and biochar – opportunities for distributed production and soil quality enhancement. In: Sustainable Alternative Fuel Feedstock Opportunities, Challenges and Roadmaps for Six U.S. regions. Proceedings of the Sustainable Feedstocks for Advanced Biofuels Workshop, Atlanta, GA: SWCS publisher, 257-291. |
| [16] | Deenik JL, Antal Jr MJ, Campbell S (2010) Charcoal volatile matter content influences plant growth and soil nitrogen transformations. Soil Fertil Plant Nutr 74: 1259-1279. |
| [17] | DeLuca TH, Mac Kenzie MD, Gundale MJ (2009) Biochar effects on soil nutrient transformation. In: Lehmann J, Joseph S, eds. Biochar for Environmental Management: Science and Technology. London: Earthscan, 251-270. |
| [18] |
Zaho L, Cao X, Masek O, et al. (2013) Heterogeneity of biochar properties as a function of feedstock sources and production temperatures. J Hazard Mater 256-257: 1-9. doi: 10.1016/j.jhazmat.2013.04.015
|
| [19] |
Major J, Rondon M, Molina D, et al. (2010) Maize yield and nutrition during 4 years after biochar application to a Colombian savanna oxisol. Plant Soil 333: 117-128. doi: 10.1007/s11104-010-0327-0
|
| [20] |
Schulz H, Glaser B (2012) Effects of biochar compared to organic and inorganic fertilizers on soil quality and plant growth in a greenhouse experiment. J Plant Nutr Soil Sci 175: 410-422. doi: 10.1002/jpln.201100143
|
| [21] |
Vaccari FP, Baronti S, Lugato E, et al. (2011) Biochar as a strategy to sequester carbon and increase yield in durum wheat. Eur J Agron 34: 231-238. doi: 10.1016/j.eja.2011.01.006
|
| [22] |
Baronti S, Vaccari FP, Miglietta F, et al. (2014) Impact of biochar application on plant water relations in Vitis vinifera (L.). Eur J Agron 53: 38-44. doi: 10.1016/j.eja.2013.11.003
|
| [23] |
Schmidt H-P, Kammann C, Niggli C, et al. (2014) Biochar and biochar-compost as soil amendments to a vineyard soil: Influences on plant growth, nutrient uptake, plant health and grape quality. Agric Ecosyst Environ 191: 117-123. doi: 10.1016/j.agee.2014.04.001
|
| [24] |
Genesio L, Miglietta F, Baronti S, et al. (2015) Biochar increases vineyard productivity without affecting grape quality: Results from a four years field experiment in Tuscany. Agr Ecos Envir 201: 20-25. doi: 10.1016/j.agee.2014.11.021
|
| [25] | Morlat R, Jacquet A (1993) The soil effects on the grapevine root system in several vineyards of the Loire Valley (France). Vitis 32: 35-42. |
| [26] | Vršič S, Kocsis L, Pulko B (2016) Influence of substrate pH on root growth, biomass and leaf mineral contents of grapevine rootstocks grown in pots. J Agr Sci Tech 18: 483-490. |
| [27] |
Zamanidis P, Paschalidis C, Maltabar L, et al. (2013) Effect of the substrates on the production of engrafted vine cuttings in heated greenhouses. Comm Soil Sci Plant Anal 44: 2488-2495. doi: 10.1080/00103624.2013.803571
|
| [28] | Conversa G, Bonasia A, Lazzizera C, et al. (2015) Influence of biochar, mycorrhizal inoculation, and fertilizer rate on growth and flowering of Pelargonium (Pelargonium zonale L.) plants. Front Plant Sci 6: 429. |
| [29] |
Graber ER, Tsechansky L, Mayzlish-Gati E, et al. (2015) A humic substances product extracted from biochar reduces Arabidopsis root hair density and length under P-sufficient and P-starvation conditions. Plant Soil 395: 21-30. doi: 10.1007/s11104-015-2524-3
|
| [30] |
Tian Y, Sun X, Li S, et al. (2012) Biochar made from green waste as peat substitute in growth media for Calathea rotundifolia cv. Fasciata. Sci Hortic-Amsterdam 143: 15-18. doi: 10.1016/j.scienta.2012.05.018
|
| [31] |
Zhang L, Sun X-Y, Tian Y, et al. (2014) Biochar and humic acids amendments improve the quality of composted green waste as a growth medium for the ornamental plant Calathea insignis. Sci Hortic 176: 70-78. doi: 10.1016/j.scienta.2014.06.021
|
| [32] |
Chan KY, Van Zwieten L, Meszaros I, et al. (2007) Agronomic values of greenwaste biochar as a soil amendment. Austral J Soil Res 45: 629-634. doi: 10.1071/SR07109
|
| [33] | Sabir A (2016) Vegetative and reproductive growth responses of grapevine cv Italia (Vitis vinifera L.) grafted on different rootstocks to contrasting soil water status. J Agr Sci Tech 18: 1681-1692. |
| [34] |
Vaughn SF, Kenar JA, Thompson AR, et al. (2013) Comparison of biochars derived from wood pellets and pelletized wheat straw as replacements for peat in potting substrates. Ind Crops Prod 51: 437-443. doi: 10.1016/j.indcrop.2013.10.010
|
| [35] |
Dumroese RK, Heiskanen J, Englund K, et al. (2011) Pelleted biochar: chemical and physical properties show potential use as a substrate in container nurseries. Biomass Bioenerg 35: 2018-2027. doi: 10.1016/j.biombioe.2011.01.053
|
| [36] | Northup J (2013) Biochar as Replacement for Perlite in Greenhouse Soilless Substrates. Graduate Theses and Dissertations, Paper 13399, Iowa State University. |
| [37] | Futsuki D, Nitta Y, Iduhara M, et al. (2014) Expression of strawberry allergen Fra a 1 gene during fruit ripening. Acta Hortic 1049: 323-328. |
| [38] | United States Department of Agriculture – Natural Resources Conservation Service (2011) Soil survey laboratory information manual. Soil Survey Investigations Report No. 45, Version 2.0. R. Burt, ed. Aqueous extraction, Method 4.3.3. USDA-NRCS, Washington, DC. 167. |
| [39] | Nelson RE (1982) Carbonate and gypsum. In: Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties. Agronomy Monograph no 9, 2nd edn. Madison, Wisc: American Society of Agronomy, 181-197. |
| [40] |
Pituello C, Francioso O, Simonetti G, et al. (2015) Characterization of chemical-physical, structural and morphological properties of biochars from biowastes produced at different temperature. J Soil Sediment 15: 792-804. doi: 10.1007/s11368-014-0964-7
|
| [41] | Cavins TJ, Whipker BE, Fonteno WC, et al. (2000) Establishing a PourThru sampling program. part 1. Ohio Florists' Assn Bul 846: 1-11. |
| [42] | Cavins TJ, Whipker BE, Fonteno WC (2004) Establishment of calibration curves for comparing pour-through and saturated media extract nutrient values. HortSci 39: 1635-1639. |
| [43] | Sokal RR, Rohlf FJ (1969) Biometry. 1st ed. San Francisco, CA, Freeman. |
| [44] |
Kaufman JH, Metin S, Saperstein DD (1989) Symmetry breaking in nitrogen-doped amorphous carbon: infrared observation of the Raman-active G and D bands. Phys Rev B 39: 13053. doi: 10.1103/PhysRevB.39.13053
|
| [45] | Giovannini C, Montecchio D, Gioacchini P, et al. (2009) Characterization of compost-based growing media. A chemical, thermal, spectroscopic and isotopic approach. Dyn Soil Dyn Plant 3: 86-92. |
| [46] | Francioso O, Ciavatta C, Tugnoli V, et al. (1998) Spectroscopic characterization of pyrophosphate incorporation during extraction of peat humic acids. Soil Sci Soc Am J 627: 181-187. |
| [47] | Novak JM, Lima I, Xing B, et al. (2009) Characterization of designer biochar produced at different temperatures and their effects on a loamy sand. Ann Environ Sci 3: 195-206. |
| [48] |
Li X, Shen Q, Zhang D, et al. (2013) Functional groups determine biochar properties (pH and EC) as studied by two-dimensional 13C NMR correlation spectroscopy. PLoS One 8: e65949. doi: 10.1371/journal.pone.0065949
|
| [49] |
Sigua GC, Novak JM, Watts DW, et al. (2016) Efficacies of designer biochars in improving biomass and nutrient uptake of winter wheat grown in a hard setting subsoil layer. Chemosphere 142: 176-183. doi: 10.1016/j.chemosphere.2015.06.015
|
| [50] |
Poorter H, Fiorani F, Stitt M, et al. (2012) The art of growing plants for experimental purposes: a practical guide for the plant biologist. Funct Plant Biol 39: 821-838. doi: 10.1071/FP12028
|
| [51] |
Vaccari FP, Maienza A, Miglietta F, et al. (2015) Biochar stimulates plant growth but not fruit yield of processing tomato in a fertile soil. Agr Ecosyst Environ 207: 163-170. doi: 10.1016/j.agee.2015.04.015
|
| [52] |
Pizzeghello D, Francioso O, Ertani A, et al. (2013) Isopentenyladenosine and cytokinin-like activity of different humic substances. J Geochem Explor 129: 70-75. doi: 10.1016/j.gexplo.2012.10.007
|
| [53] |
Jindo K, Sonoki T, Matsumoto K, et al. (2016) Influence of biochar addition on the humic substances of composting manures. Waste Manage 49: 545-552. doi: 10.1016/j.wasman.2016.01.007
|
| [54] |
Nardi S, Pizzeghello D, Schiavon M, et al. (2016) Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci Agr 73: 18-23. doi: 10.1590/0103-9016-2015-0006
|
| [55] | Nardi S, Carletti P, Pizzeghello D, et al. (2009) Biological activities of humic substances. In: Senesi N, Xing B, Huang PM, eds. Biophysico-Chemical Processes Involving Natural Nonliving Organic Matter in Environmental Systems. PART I. Fundamentals and impact of mineral-organic-biota interactions on the formation, transformation, turnover, and storage of natural nonliving organic matter (NOM), Volume 2, Hoboken, NJ: John Wiley & Sons, 305-340. |
| [56] |
Dana EM, Juan JG (2004) Distortion of the SPAD 502 chlorophyll meter readings by changes in irradiance and leaf water status. Agronomie 24: 41-46. doi: 10.1051/agro:2003060
|
| [57] |
Chalker-Scott L (1999) Environmental significance of anthocyanins in plant stress responses. Photochem Photobiol 70: 1-9. doi: 10.1111/j.1751-1097.1999.tb01944.x
|
| [58] |
Kuzyakov Y, Subbotina I, Chen H, et al. (2009) Black carbon decomposition and incorporation into soil microbial biomass estimated by 14C labeling. Soil Biol Biochem 41: 210-219. doi: 10.1016/j.soilbio.2008.10.016
|

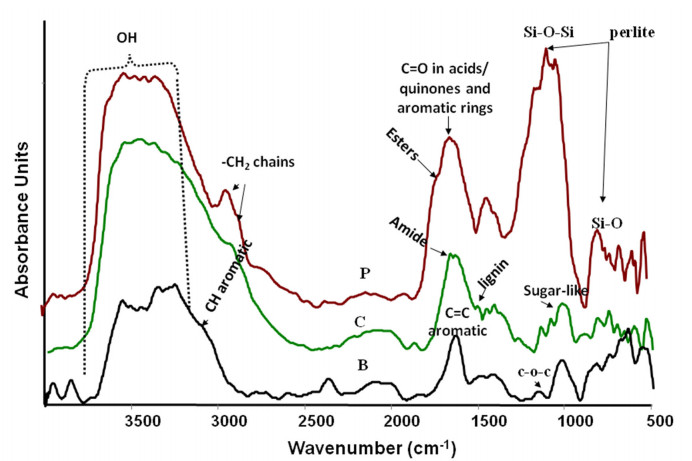
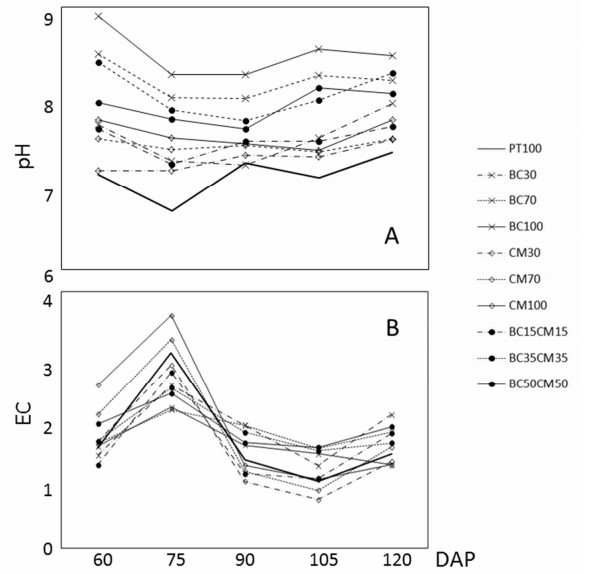
Figures(3) / Tables(5)

Arianna Bozzolo, Diego Pizzeghello, Alessandra Cardinali, Ornella Francioso, Serenella Nardi. Effects of moderate and high rates of biochar and compost on grapevine growth in a greenhouse experiment[J]. AIMS Agriculture and Food, 2017, 2(1): 113-128. doi: 10.3934/agrfood.2017.1.113







 DownLoad:
DownLoad: