Citation: Paul Agu Igwe, Mahfuzur Rahman, Kenny Odunukan, Nonso Ochinanwata, Obiamaka P. Egbo, Chinedu Ochinanwata. Drivers of diversification and pluriactivity among smallholder farmers—evidence from Nigeria[J]. Green Finance, 2020, 2(3): 263-283. doi: 10.3934/GF.2020015

| [1] |
Abdulai A, CroleRees A (2001) Determinants of Income Diversification amongst Rural Households in Southern Mali. Food Policy 26: 437-452. doi: 10.1016/S0306-9192(01)00013-6

|
| [2] |
Akinrinde AF, Omotesho KF, Ogulande I (2018) The issue of income diversification among rural farming households: empirical evidence from Kwara State, Nigeria. J Agribus Rural Dev 49: 231-238. doi: 10.17306/J.JARD.2018.00427
|
| [3] |
Atuoye KN, Antabe R, Sano Y, et al. (2019) Household income diversification and food insecurity in the upper west region of Ghana. Social Indic Res 144: 899-920. doi: 10.1007/s11205-019-02062-7
|
| [4] |
Bezu S, Barrett C (2012) Employment dynamics in the rural nonfarm sector in Ethiopia: Do the poor have time on their side? J Dev Stud 48: 1223-1240. doi: 10.1080/00220388.2012.671476
|
| [5] | Carr S (2016) An exploration of Agripreneurship, Scope, Actors and Prospects. Available from: https://cdn.ymaws.com/www.andeglobal.org/resource/dynamic/blogs/20160701_143408_24248.pdf. |
| [6] | Chambers R, Conway GR (1992) Sustainable rural livelihoods: Practical concepts for the 21st century. IDS Discussion Paper No. 296. IDS, Brighton, UK. |
| [7] | Chandio AA, Jiang Y, Wei F, et al. (2017) Famers' access to credit: Does collateral matter or cash flow matter?-Evidence from Sindh, Pakistan. Cogent Econ Financ 5: 1-17. |
| [8] | CIA World Factbook (2016) Eastern-Nigerian population. Center for the Study of Intelligence, USA. Available from: www.cia.gov/library/publications/resources/the-world-factbook/geos/rp.html. |
| [9] |
Deininger K, Savastano S, Xia F (2017) Smallholders' land access in Sub-Saharan Africa: A new landscape? Food policy 67: 78-92. doi: 10.1016/j.foodpol.2016.09.012
|
| [10] |
Dessart FJ, Barreiro-Hurlé J, van Bavel R (2019) Behavioural factors affecting the adoption of sustainable farming practices: a policy-oriented review. Eur Rev Agr Econ 46: 417-471. doi: 10.1093/erae/jbz019
|
| [11] |
Edelman LF, Brush CG, Manolova TS, et al. (2010) Start-up Motivations and Growth Intentions of Minority Nascent Entrepreneurs. J Small Bus Manage 48: 174-196. doi: 10.1111/j.1540-627X.2010.00291.x
|
| [12] | Ekekwe N (2017) How Digital Technology Is Changing Farming in Africa. Harvard Bus Rev 18. |
| [13] | Ezeakor A (2011) All Alone, the Psychosocial Condition of Nigerian Widows and Childless Women: Implication for Survivors' Cases, Xlibris Publishing, New York, USA. |
| [14] |
Fabusoro E, Omotayo A, Apantaku S, et al. (2010) Forms and determinants of rural livelihoods diversification in Ogun State, Nigeria. J Sustain Agr 34: 417-438. doi: 10.1080/10440041003680296
|
| [15] | FAO (2020) Nigeria at a glance. Rome, Italy. Available from: http://www.fao.org/nigeria/fao-in-nigeria/nigeria-at-a-glance/en/. |
| [16] |
Gautam Y, Andersen P (2017) Rural livelihood diversification and household well-being: Insights from Humla, Nepal. J Rural Stud 44: 239-249. doi: 10.1016/j.jrurstud.2016.02.001
|
| [17] |
Gedajlovic E, Honig B, Moore CB, et al. (2013) Social Capital and Entrepreneurship: A Schema and Research Agenda. Entrep Theory Pract 37: 455-478. doi: 10.1111/etap.12042
|
| [18] | Getu M, Devereux S (2013) Informal and formal Protection Systems in Sub-Saharan Africa, Fountain Publishers, Kampala, Ethiopia. |
| [19] |
Haggblade S, Hazell P, Reardon T (2010) The Rural Nonfarm Economy: Prospects for Growth and Poverty Reduction. World Dev 38: 1429-1441. doi: 10.1016/j.worlddev.2009.06.008
|
| [20] | Igwe PA, Amaugo AN, Ogundana OM, et al. (2018) Factors affecting the investment climate, SMEs productivity and entrepreneurship in Nigeria. Eur J Sust Dev 7: 1-19. |
| [21] |
Igwe PA, Madichie NO, Newbery R (2019) Determinants of livelihood choices and artisanal entrepreneurship in Nigeria. Int J Entrep Beha Res 25: 674-697. doi: 10.1108/IJEBR-02-2018-0102
|
| [22] |
Jackson C (2003) Gender Analysis of Land: Beyond Land Rights for Women? J Agrar Change 3: 453-480. doi: 10.1111/1471-0366.00062
|
| [23] |
Jansen HGP, Pender J, Damon A, et al. (2006) Policies for sustainable development in the hillside areas of Honduras: quantitative livelihoods approach. Agr Econ 34: 141-153. doi: 10.1111/j.1574-0864.2006.00114.x
|
| [24] |
Kandker SR, Koolwall GB (2010) How Infrastructure and Financial Institutions Affect Rural Income and Poverty: Evidence from Bangladesh. J Dev Stud 46: 1109-1129. doi: 10.1080/00220380903108330
|
| [25] | Kim PH, Aldrich HE (2006) Social Capital and Entrepreneurship, Now Publishers Inc. |
| [26] |
Kwon SW, Heflin C, Ruef M (2013) Community social capital and entrepreneurship. Am Sociol Rev 78: 980-1008. doi: 10.1177/0003122413506440
|
| [27] |
Kimmitt J, Muñoz P, Newbery R (2019) Poverty and the varieties of entrepreneurship in the pursuit of prosperity. J Bus Ventur 35: 105939. doi: 10.1016/j.jbusvent.2019.05.003
|
| [28] |
Kouamé A, Schellekens J (2002) Développement rural et attitude à l'égard de la taille de la famille en Côte d'Ivoire. Population 57: 269-299. doi: 10.3917/pope.202.0269
|
| [29] | Kraaijenbrink J, Spender JC, Groen AJ (2010) The Resource-Based View: A Review and Assessment of Its Critiques. J Manage 36: 349-372. |
| [30] |
Kuivanen KS, Michalscheck M, Descheemaeker K, et al. (2016) A comparison of statistical and participatory clustering of smallholder farming systems: a case study in Northern Ghana. J Rural Stud 45:184-198. doi: 10.1016/j.jrurstud.2016.03.015
|
| [31] |
Loison SA (2015) Rural Livelihood Diversification in Sub-Saharan Africa: A Literature Review. J Dev Stud 51: 1125-1138. doi: 10.1080/00220388.2015.1046445
|
| [32] |
Morgan SL, Marsden T, Miele M, et al. (2010) Agricultural multi-functionality and farmers' entrepreneurial skills: a study of Tuscan and Welsh farmers. J Rural Stud 26: 116-129. doi: 10.1016/j.jrurstud.2009.09.002
|
| [33] | Morse S, McNamara N, Acholo M (2009) Sustainable Livelihood Approach: A critical analysis of theory and practice. Geographical Paper No. 189. Accesses 10/04/2019, Available from: https://www.reading.ac.uk/web/files/geographyandenvironmentalscience/GP189.pdf. |
| [34] |
Morris W, Henley A, Dowell D (2017) Farm diversification, entrepreneurship and technology adoption: Analysis of upland farmers in Wales. J Rural Stud 53: 132-143. doi: 10.1016/j.jrurstud.2017.05.014
|
| [35] |
Nagler P, Naudé W (2017) Non-farm entrepreneurship in rural sub-Saharan Africa: New empirical evidence. Food Policy 67: 175-191. doi: 10.1016/j.foodpol.2016.09.019
|
| [36] | Nagler P, Naude W (2018) Nonfarm Enterprises in Rural Africa Chapter 9, In: Christiaensen Luc, and Lionel Demery, eds., Agriculture in Africa: Telling Myths from Facts, Directions in Development, Washington, DC: World Bank. |
| [37] | Naminse EY, Zhuang J (2018) Does farmer entrepreneurship alleviate rural poverty in China? Evidence from Guangxi Province. PLoS ONE 13: 1-18. |
| [38] |
Ndaeyo NU, Umoh GS, Ekpe EO (2001) Farming Systems in South-eastern Nigeria: Implications for Sustainable Agricultural Production. J Sustain Agr 17: 75-89. doi: 10.1300/J064v17n04_07
|
| [39] | Nigerian National Agricultural Sample Survey NASS (2010/2011) National Bureau of Statistics (NBS) and Federal Ministry of Agriculture and Rural Development, Abuja. Available from: https://www.nigerianstat.gov.ng/nada/index.php/catalog/31. |
| [40] | Nwokoro CV, Chima FO (2017) Impact of Environmental Degradation on Agricultural Production and Poverty in Rural Nigeria. Am Int J Contemp Res 7: 6-14. |
| [41] |
Ochieng J, Knerr B, Owuor G, et al. (2019) Food crops commercialization and household livelihoods: Evidence from rural regions in Central Africa. Agribusiness 36: 318-338. doi: 10.1002/agr.21619
|
| [42] |
Ola O, Menapace L (2020) Revisiting constraints to smallholder participation in high-value markets: A best-worst scaling approach. Agr Econ 51: 595-608. doi: 10.1111/agec.12574
|
| [43] | Onyeiwu S, Liu J (2011) Determinants of Income Poverty in Rural Africa: Empirical Evidence from Kenya and Nigeria. In African Economic Conference, Addis Ababa, 26-28. |
| [44] |
Partanen J, Kohtamäki M, Parida V, et al. (2017) Developing and validating a multi-dimensional scale for operationalizing industrial service offering. J Bus Ind Mark 32: 295-309. doi: 10.1108/JBIM-08-2016-0178
|
| [45] |
Pindado E, Sánchez M (2018) Growth-oriented new agricultural ventures: the role of entrepreneurial resources and capabilities under convergence forces. Eur Rev Agr Econ 46: 800-833. doi: 10.1093/erae/jby039
|
| [46] |
Radicic D, Bennett R, Newton G (2017) Portfolio entrepreneurship in farming: Empirical evidence from the 1881 Census for England and Wales. J Rural Stud 55: 289-302. doi: 10.1016/j.jrurstud.2017.08.019
|
| [47] |
Rahman S, Akter S (2014) Determinants of livelihood choices: an empirical analysis from rural Bangladesh. J S Asian Dev 9: 287-308. doi: 10.1177/0973174114549101
|
| [48] |
Rahman M, Akter M, Radicic D (2020) Internationalization as a strategy for small and medium-sized enterprises and the impact of regulatory environment: An emerging country perspective. Bus Strat Dev 3: 213-225. doi: 10.1002/bsd2.90
|
| [49] |
Rahut DB, Scharf MM (2012) Livelihood diversification strategies in the Himalayas. Aust J Agr Resour Econ 56: 1-19. doi: 10.1111/j.1467-8489.2011.00567.x
|
| [50] |
Sanou A, Liverpool-Tasie LSO, Kerr JM (2020) Threshold investment and firm viability: Evidence from commercial poultry farms in Nigeria. Agribusiness 36: 422-439. doi: 10.1002/agr.21639
|
| [51] |
Sarker MNI, Wu M, Alam GM, et al. (2020) Livelihood diversification in rural Bangladesh: Patterns and determinants in disaster prone riverine islands. Land Use Policy 96: 104720. doi: 10.1016/j.landusepol.2020.104720
|
| [52] |
Sarker MNI, Wu M, Alam GM, et al. (2020) Livelihood resilience of riverine island dwellers in the face of natural disasters: Empirical evidence from Bangladesh. Land Use Policy 95: 104599. doi: 10.1016/j.landusepol.2020.104599
|
| [53] | Sarker MNI, Wu M, Alam GM, et al. (2019) Livelihood vulnerability of riverine-island dwellers in the face of natural disasters in Bangladesh. Sustainability 11: 16-23. |
| [54] | Sen A (1985) Commodities and Capabilities, Amsterdam: North-Holland. |
| [55] | United Nations (UN) About the Sustainable Development Goals. Available from: https://www.un.org/sustainabledevelopment/sustainable-development-goals/. |
| [56] | Uzuegbunam AO, Uzuegbunam I (2018) Arm's-length or give-and-take? Gender differences in the relational orientation of new ventures in Sub-Saharan Africa. Strat Entrep J 12: 522-541. |
| [57] |
Wambua S, Birachi E, Gichangi A, et al. (2018) Influence of productive resources on bean production in male- and female-headed households in selected bean corridors of Kenya. Agr Food Secur 7: 85-112. doi: 10.1186/s40066-018-0236-7
|
| [58] | Wardhana D, Ihle R, Heijman W (2020) Farmer cooperation in agro-clusters: Evidence from Indonesia. Agribusiness, 1-26. |
| [59] | World Development Report (2008) Agriculture for Development, The World Bank, Washington, DC: USA. |
| [60] | World Bank (2018) In Africa, more not fewer people will work in agriculture. Available from: https://blogs.worldbank.org/jobs/africa-more-not-fewer-people-will-work-agriculture. |
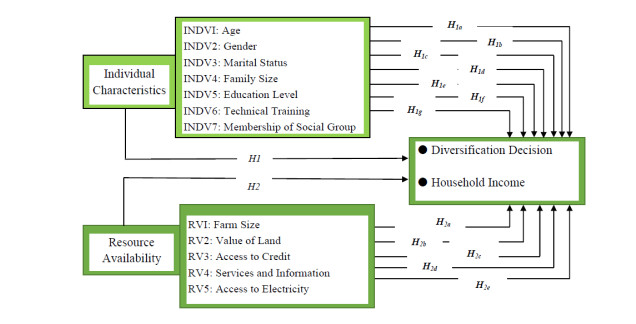
Figures(1) / Tables(5)

Paul Agu Igwe, Mahfuzur Rahman, Kenny Odunukan, Nonso Ochinanwata, Obiamaka P. Egbo, Chinedu Ochinanwata. Drivers of diversification and pluriactivity among smallholder farmers—evidence from Nigeria[J]. Green Finance, 2020, 2(3): 263-283. doi: 10.3934/GF.2020015







 DownLoad:
DownLoad: