Citation: Nataša Čerekovic, Palmiro Poltronieri. Plant signaling pathways activating defence response and interfering mechanisms by pathogen effectors, protein decoys and bodyguards[J]. AIMS Molecular Science, 2017, 4(3): 370-388. doi: 10.3934/molsci.2017.3.370

| [1] |
Erbs G, Silipo A, Aslam S, et al. (2008) Peptidoglycan and muropeptides from pathogens Agrobacterium and Xanthomonas elicit plant innate immunity: structure and activity. Chem Biol 15: 438-448. doi: 10.1016/j.chembiol.2008.03.017

|
| [2] |
Ranf S, Gisch N, Schaffer M, et al. (2015) A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nature Immunol 16: 426-433. doi: 10.1038/ni.3124
|
| [3] |
Cook DE, Mesarich CH, Thomma BP (2015) Understanding plant immunity as a surveillance system to detect invasion. Annu Rev Phytopathol 53: 541-563. doi: 10.1146/annurev-phyto-080614-120114
|
| [4] |
Klarzynski O, Plesse B, Joubert JM, et al.(2000) Linear beta-1,3 glucans are elicitors of defense responses in tobacco. Plant Physiol 124: 1027-1038. doi: 10.1104/pp.124.3.1027
|
| [5] |
Monaghan J, Zipfel C (2012) Plant pattern recognition receptor complexes at the plasma membrane. Curr Opin Plant Biol 15: 349-357. doi: 10.1016/j.pbi.2012.05.006
|
| [6] |
Zhang L, Kars I, Essenstam B, et al. (2014) Fungal endopolygalacturonases are recognized as microbe-associated molecular patterns by the Arabidopsis receptor-like protein RESPONSIVENESS TO BOTRYTIS POLYGALACTURONASES1. Plant Physiol 164: 352-364. doi: 10.1104/pp.113.230698
|
| [7] |
Jehle AK, Fürst U, Lipschis M, et al. (2013) Perception of the novel MAMP eMax from different Xanthomonas species requires the Arabidopsis receptor-like protein ReMAX and the receptor kinase SOBIR. Plant Signal Behav 8: e27408. doi: 10.4161/psb.27408
|
| [8] |
Zipfel C, Robatzek S, Navarro L, et al. (2004) Bacterial disease resistance in Arabidopsis through flagellin perception. Nature 428: 764-767. doi: 10.1038/nature02485
|
| [9] |
Felix G, Duran JD, Volko S, et al. (1999) Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J 18: 265-276. doi: 10.1046/j.1365-313X.1999.00265.x
|
| [10] | Smith KD, Andersen-Nissen E, Hayashi F, et al. (2003) Toll-like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nature Immunol 4: 1247-1253. |
| [11] |
Oome S, Raaymakers TM, Cabral A, et al. (2014) Nep1-like proteins from three kingdoms of life act as a microbe-associated molecular pattern in Arabidopsis. Proc Natl Acad Sci USA 111: 16955-16960. doi: 10.1073/pnas.1410031111
|
| [12] |
Granato D, Bergonzelli GE, Pridmore RD, et al. (2004) Cell surface-associated elongation factor Tu mediates the attachment of Lactobacillus johnsonii NCC533 (La1) to human intestinal cells and mucins. Infect Immun 72: 2160-2169. doi: 10.1128/IAI.72.4.2160-2169.2004
|
| [13] |
Jonák J (2007) Bacterial elongation factors EF-Tu, their mutants, chimeric forms, and domains: Isolation and purification. J Chromatogr B 849: 141-153. doi: 10.1016/j.jchromb.2006.11.053
|
| [14] |
Zipfel C, Kunze G, Chinchilla D, et al. (2006) Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell 125: 749-760. doi: 10.1016/j.cell.2006.03.037
|
| [15] |
Kunze G, Zipfel C, Robatzek S, et al. (2004) The N terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. Plant Cell 16: 3496-3507. doi: 10.1105/tpc.104.026765
|
| [16] |
Lacombe S, Rougon-Cardoso A, Sherwood E, et al. (2010) Interfamily transfer of a plant pattern-recognition receptor confer broad-spectrum bacterial resistance. Nat Biotechnol 28: 365-369. doi: 10.1038/nbt.1613
|
| [17] |
Schwessinger B, Bahar O, Thomas N, et al. (2015) Transgenic Expression of the Dicotyledonous Pattern Recognition Receptor EFR in Rice Leads to Ligand-Dependent Activation of Defense Responses. PLoS Pathog 11: e1004809. doi: 10.1371/journal.ppat.1004809
|
| [18] | Furukawa T, Inagaki H, Takai R, et al. (2013) Two distinct EF-Tu epitopes induce immune responses in rice and Arabidopsis. Mol Plant-Microbe Interact 27: 113-124. |
| [19] |
Hind SR, Strickler SR, Boyle PC, et al. (2016) Tomato receptor FLAGELLIN-SENSING 3 binds flgII-28 and activates the plant immune system. Nat Plants 2: 16128. doi: 10.1038/nplants.2016.128
|
| [20] |
Katsuragi Y, Takai R, Furukawa T, et al. (2015) C-terminal is recognised by a different flagellin sensing receptor. Mol Plant-Microbe Interact 28: 648-658. doi: 10.1094/MPMI-11-14-0372-R
|
| [21] |
Hirai H, Takai R, Iwano M, et al. (2011) Glycosylation regulates specific induction of rice immune responses by Acidovorax avenae flagellin. J Biol Chem 286: 25519-25530. doi: 10.1074/jbc.M111.254029
|
| [22] |
Hirai H, Takai R, Kondo M, et al. (2014) Glycan moiety of flagellin in Acidovorax avenae K1 prevents the recognition by rice that causes the induction of immune responses. Plant Signal Behav 9: e972782. doi: 10.4161/psb.29933
|
| [23] |
Takeuchi K, Ono H, Yoshida M, et al. (2007) Flagellin glycans from two pathovars of Pseudomonas syringae contain rhamnose in D and L configurations in different ratios and modified 4-amino-4,6-dideoxyglucose. J Bacteriol 189: 6945-56. doi: 10.1128/JB.00500-07
|
| [24] |
Che FS, Nakajima Y, Tanaka N, et al. (2000) Flagellin from an incompatible strain of Pseudomonas avenae induces a resistance response in cultured rice cells. J Biol Chem 275: 32347-32356. doi: 10.1074/jbc.M004796200
|
| [25] |
Taguchi F, Shimizu R, Inagaki Y, et al. (2003) Post-translational modification of flagellin determines the specificity of HR induction. Plant Cell Physiol 44: 342-349. doi: 10.1093/pcp/pcg042
|
| [26] |
Yamamoto M, Ohnishi-Kameyama M, Nguyen CL, et al. (2011) Identification of genes involved in the glycosylation of modified viosamine of flagellins in Pseudomonas syringae by mass spectrometry. Genes 2: 788-803. doi: 10.3390/genes2040788
|
| [27] |
Sun Y, Li L, Macho AP, et al. (2013) Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science 342: 624-628. doi: 10.1126/science.1243825
|
| [28] |
Sun W, Dunning FM, Pfund C, et al. (2006) Within-Species flagellin polymorphism in Xanthomonas campestris pv campestris and its impact on elicitation of Arabidopsis FLAGELLIN SENSING2–dependent defenses. Plant Cell 18: 764-779. doi: 10.1105/tpc.105.037648
|
| [29] |
Pel MJC, van Dijken AJH, Bardoel BW, et al. (2014) Pseudomonas syringae evades host immunity by degrading flagellin monomers with alkaline protease AprA. Mol Plant-Microbe Interact 27: 603-610. doi: 10.1094/MPMI-02-14-0032-R
|
| [30] |
Eckardt NA (2017) The plant cell reviews plant immunity: Receptor-Like Kinases, ROS-RLK crosstalk, quantitative resistance, and the growth/defense trade-off. Plant Cell 29: 601-602. doi: 10.1105/tpc.17.00289
|
| [31] |
Bi G, Liebrand TW, Bye RR, et al. (2016) SOBIR1 requires the GxxxG dimerization motif in its transmembrane domain to form constitutive complexes with receptor-like proteins. Mol Plant Pathol 17: 96-107. doi: 10.1111/mpp.12266
|
| [32] |
Albert I, Böhm H, Albert M, et al. (2015) An RLP23–SOBIR1–BAK1 complex mediates NLP-triggered immunity. Nature Plants 1: 15140. doi: 10.1038/nplants.2015.140
|
| [33] |
Heese A, Hann DR, Gimenez-Ibanez S, et al. (2007) The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc Natl Acad Sci USA 104: 12217-12222. doi: 10.1073/pnas.0705306104
|
| [34] |
Chinchilla D, Zipfel C, Robatzek S, et al. (2007) A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 448: 497-500. doi: 10.1038/nature05999
|
| [35] |
Schwessinger B, Roux M, Kadota Y, et al. (2011) Phosphorylation-dependent differential regulation of plant growth, cell death, and innate immunity by the regulatory receptor-like kinase BAK1. PLoS Genet 7: e1002046. doi: 10.1371/journal.pgen.1002046
|
| [36] |
Somssich M, Ma Q, Weidtkamp-Peters S, et al. (2015) Real-time dynamics of peptide ligand-dependent receptor complex formation in planta. Sci Signal 8: ra76. doi: 10.1126/scisignal.aab0598
|
| [37] | Dufayard JF, Bettembourg M, Fischer I, et al. (2017) New insights on Leucine-Rich Repeats Receptor-like kinase orthologous relationships in Angiosperms. Front Plant Sci 8: 381. |
| [38] |
Tang D, Wang G, Zhou JM (2017) Receptor kinases in plant-pathogen interactions: More than pattern recognition. Plant Cell 29: 618-637. doi: 10.1105/tpc.16.00891
|
| [39] |
Bigeard J, Colcombet J, Hirt H (2015) Signaling mechanisms in Pattern-Triggered Immunity (PTI). Mol Plant 8: 521-539. doi: 10.1016/j.molp.2014.12.022
|
| [40] |
Wu S, Shan L, He P (2014) Microbial signature-triggered plant defense responses and early signaling mechanisms. Plant Sci 228: 118-126. doi: 10.1016/j.plantsci.2014.03.001
|
| [41] |
Song WY, Wang GL, Chen LL, et al. (1995) A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 270: 1804-1806. doi: 10.1126/science.270.5243.1804
|
| [42] |
Kimura S, Waszczak C, Hunter K, et al. (2017) Bound by fate: the role of reactive oxygen species in receptor-like kinase signaling. Plant Cell 29: 638-654. doi: 10.1105/tpc.16.00947
|
| [43] |
Karasov TL, Chae E, Herman JJ, et al. (2017) Mechanisms to mitigate the trade-off between growth and defense. Plant Cell 29: 666-680. doi: 10.1105/tpc.16.00931
|
| [44] |
Santino A, Taurino M, De Domenico S, et al. (2013) Jasmonate signalling in plant defense response to multiple (a)biotic stresses. Plant Cell Rep 32: 1085-1098. doi: 10.1007/s00299-013-1441-2
|
| [45] | Poltronieri P, Taurino M, Bonsegna S, et al. (2014) Nitric Oxide: Detection Methods and Possible Roles During Jasmonate-regulated Stress Response, In: Khan MN, Mobin M, Mohammad F, Corpas FJ, Authors, Nitric Oxide in Plants: Metabolism and Role in Stress Physiology, Switzerland: Springer, 127-138. |
| [46] | Santino A, Bonsegna S, De Domenico S, et al. (2010) Plant oxylipins and their contribution to plant defence. Curr Topics Plant Biol 11: 103-111 |
| [47] | Klessig DF, Tian M, Choi HW (2016) Multiple targets of salicylic acid and its derivatives in plants and animals. Front immunol 7: 206. |
| [48] |
Kay S, Bonas U (2009) How Xanthomonas type III effectors manipulate the host plant. Current Opin. Microbiol 12: 37-43. doi: 10.1016/j.mib.2008.12.006
|
| [49] |
Lee S, Yang DS, Uppalapati SR, et al. (2013) Suppression of plant defense responses by extracellular metabolites from Pseudomonas syringae pv. tabaci in Nicotiana benthamiana. BMC Plant Biol 13: 65. doi: 10.1186/1471-2229-13-65
|
| [50] |
Boch J, Bonas U (2010) Xanthomonas AvrBs3 family-type III effectors: Discovery and function. Annu Rev Physiopathol 48: 419-436. doi: 10.1146/annurev-phyto-080508-081936
|
| [51] |
Macho AP, Schwessinger B, Ntoukakis V, et al. (2014) A bacterial tyrosine phosphatase inhibits plant pattern recognition receptor activation. Science 343: 1509-1512. doi: 10.1126/science.1248849
|
| [52] | Brutus A, Reca IB, Herga S, et al. (2005) A family 11 xylanase from the pathogen Botrytis cinerea is inhibited by plant endoxylanase inhibitors XIP-I and TAXI-I. Biochem Biophys Res Commun 337: 160-166. |
| [53] | Tyler BM, Rouxel T (2012) Effectors of fungi and oomycetes: their virulence and avirulence functions and translocation from pathogen to host cells. Mol Plant Immun 123-167. |
| [54] |
Macho AP, Zipfel C (2015) Targeting of plant pattern recognition receptor triggered immunity by bacterial type-III secretion system effectors. Curr Opin Microbiol 23: 14-22. doi: 10.1016/j.mib.2014.10.009
|
| [55] | Tian M, Win J, Song J, et al. (2007) A Phytophthora infestans cystatin-like protein targets a novel tomato papain-like apoplastic protease. Plant Physiol 143: 364-377. |
| [56] |
Song J, Win J, Tian M, et al. (2009) Apoplastic effectors secreted by two unrelated eukaryotic plant pathogens target the tomato defense protease Rcr3. Proc Natl Acad Sci USA 106: 1654-1659. doi: 10.1073/pnas.0809201106
|
| [57] |
Catanzariti AM, Dodds PN, Lawrence GJ, et al. (2006) Haustorially expressed secreted proteins from flax rust are highly enriched for avirulence elicitors. Plant Cell 18: 243-256. doi: 10.1105/tpc.105.035980
|
| [58] |
Tian M, Benedetti B, Kamoun S (2005) A second Kazal-like protease inhibitor from Phytophthora infestans inhibits and interacts with the apoplastic pathogenesis-related protease P69b of tomato. Plant Physiol 138: 1785-1793. doi: 10.1104/pp.105.061226
|
| [59] |
Shindo T, Misas-Villamil JC, Hörger AC, et al. (2012) A role in immunity for Arabidopsis cysteine protease RD21, the ortholog of the tomato immune protease C14. PLoS One 7: e29317. doi: 10.1371/journal.pone.0029317
|
| [60] | Jashni KM, Mehrabi R, Collemare J, et al. (2015) The battle in the apoplast: further insights in the roles of proteases and their inhibitors in plant-pathogen interactions. Front Plant Sci 6: 584. |
| [61] |
van den Hooven HW, van den Burg HA, Vossen P, et al. (2001) Disulfide bond structure of the AVR9 elicitor of the fungal tomato pathogen Cladosporium fulvum: evidence for a cystine knot. Biochem 40: 3458-3466. doi: 10.1021/bi0023089
|
| [62] |
Pogány M, Dankó T, Kámán-Tóth E (2015) Regulatory proteolysis in Arabidopsis-pathogen interactions. Int J Mol Sci 16: 23177-23194. doi: 10.3390/ijms161023177
|
| [63] |
Poltronieri P, Liu S, Cimaglia F, et al. (2012) Characterization of Kunitz-type inhibitor B1 performance using protein chips and AFM. Sensors Actuators B Chem 168: 231-237. doi: 10.1016/j.snb.2012.04.013
|
| [64] | Valueva TA, Revina TA, Mosolov VV, et al. (2000) Primary structure of potato Kunitz-type serine proteinase inhibitor. Biol Chem 381: 1215-1221. |
| [65] |
Wang W, Cai J, Wang P, et al. (2017) Post-transcriptional regulation of fruit ripening and disease resistance in tomato by the vacuolar protease SlVPE3. Genome Biology 18: 47. doi: 10.1186/s13059-017-1178-2
|
| [66] |
Jones JD, Dangl JL (2006) The plant immune system. Nature 444: 323-329. doi: 10.1038/nature05286
|
| [67] |
Ilyas M, Hörger AC, Bozkurt TO, et al. (2015) Functional divergence of two secreted immune proteases of tomato. Curr Biol 25: 2300-2306. doi: 10.1016/j.cub.2015.07.030
|
| [68] |
Houterman PM, Ma L, van Ooijen G, et al (2009) The effector protein Avr2 of the xylem-colonizing fungus Fusarium oxysporum activates the tomato resistance protein I-2 intracellularly. Plant J 58: 970-978. doi: 10.1111/j.1365-313X.2009.03838.x
|
| [69] |
Ma L, Houterman PM, Gawehns F, et al. (2015) The AVR2-SIX5 gene pair is required to activate I-2-mediated immunity in tomato. New Phytol 208: 507-518. doi: 10.1111/nph.13455
|
| [70] |
Shabab M, Shindo T, Gu C, et al. (2008) Fungal effector protein AVR2 targets diversifying defense-related cys proteases of tomato. Plant Cell 20: 1169-1183. doi: 10.1105/tpc.107.056325
|
| [71] |
López-Solanilla E, Bronstein PA, Schneider AR (2004) HopPtoN is a Pseudomonas syringae Hrp (type III secretion system) cysteine protease effector that suppresses pathogen-induced necrosis associated with both compatible and incompatible plant interactions. Mol Microbiol 54: 353-365 doi: 10.1111/j.1365-2958.2004.04285.x
|
| [72] |
Wulff BBH, Chakrabarti A, Jones DA (2009) Recognitional specificity and evolution in the tomato–Cladosporium fulvum pathosystem. Microbiol Plant Mol Int 22: 1191-1202. doi: 10.1094/MPMI-22-10-1191
|
| [73] | Lee S, Yang DS, Uppalapati SR (2013) Suppression of plant defense responses by extracellular metabolites from Pseudomonas syringae pv. tabaci in Nicotiana benthamiana. BMC Plant Biol 13: 65. |
| [74] |
Buttner D (2016) Behind the lines–actions of bacterial type III effector proteins in plant cells. FEMS Microbiol Rev 40: 894-937. doi: 10.1093/femsre/fuw026
|
| [75] | Thieme F, Szczesny R, Urban A, et al. (2007) New type III effectors from Xanthomonas campestris pv. vesicatoria trigger plant reactions dependent on a conserved N-myristoylation motif. Mol Plant-Microbe Interact 20: 1250-1261. |
| [76] |
Seto D, Koulena N, Lo T, et al. (2017) Expanded type III effector recognition by the ZAR1 NLR protein using ZED1-related kinases. Nature Plants 3: 17027. doi: 10.1038/nplants.2017.27
|
| [77] |
Lewis JD, Lo T, Bastedo P, et al. (2014) The rise of the undead: pseudokinases as mediators of effector-triggered immunity. Plant Signal Behav 9: e27563. doi: 10.4161/psb.27563
|
| [78] |
Wang G, Roux B, Feng F, et al. (2015) The decoy substrate of a pathogen effector and a pseudokinase specify pathogen-induced modified-self recognition and immunity in plants. Cell Host Microbe 18: 285-295. doi: 10.1016/j.chom.2015.08.004
|
| [79] |
Baudin M, Hassan JA, Schreiber KJ, et al. (2017) Analysis of the ZAR1 immune complex reveals determinants for immunity and molecular interactions. Plant Physiol 174: 2038-2053. doi: 10.1104/pp.17.00441
|
| [80] |
Trda L, Fernandez O, Boutrot F, et al. (2014) The grapevine flagellin receptor VvFLS2 differentially recognizes flagellin-derived epitopes from the endophytic growth promoting bacterium Burkholderia phytofirmans and plant pathogenic bacteria. New Phytol 201: 1371-1384. doi: 10.1111/nph.12592
|
| [81] |
Zhang X, Dong W, Sun J, et al. (2015) The receptor kinase CERK1 has dual functions in symbiosis and immunity signalling. Plant J 81: 258-267. doi: 10.1111/tpj.12723
|
| [82] | Zeilinger S, Gupta VK, Dahms TE, et al. (2016) Friends or foes? Emerging insights from fungal interactions with plants. FEMS Microbiol Rev 40: 182-207. |
| [83] | Genre A, Russo G (2016) Does a common pathway transduce symbiotic signals in plant-microbe interactions? Front Plant Sci 7: 96. |
| [84] |
Rasmussen SR, Füchtbauer W, Novero M, et al. (2016) Intraradical colonization by arbuscular mycorrhizal fungi triggers induction of a lipochitooligosaccharide receptor. Sci Rep 6: 29733. doi: 10.1038/srep29733
|
| [85] |
Akamatsu A, Shimamoto K, Kawano Y (2016) Crosstalk of signaling mechanisms involved in host defense and symbiosis against microorganisms in rice. Curr Genomics 17: 297-307. doi: 10.2174/1389202917666160331201602
|
| [86] |
Shinya T, Nakagawa T, Kaku H, et al. (2015) Chitin-mediated plant-fungal interactions: catching, hiding and handshaking. Curr Opin Plant Biol 26: 64-71. doi: 10.1016/j.pbi.2015.05.032
|
| [87] | Ma Z, Zhu L, Song T, et al. (2017) A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor. Science 355: 710-714. |
| [88] | Paulus JK, Kourelis J, van der Hoorn RAL (2017) Bodyguards: Pathogen-derived decoys that protect virulence factors. Trends Plant Sci 22: 355-357. |
| [89] |
Ji Z, Ji C, Liu B, et al. (2016) Interfering TAL effectors of Xanthomonas oryzae neutralize R-gene-mediated plant disease resistance. Nature Comm 7: 13435. doi: 10.1038/ncomms13435
|
| [90] |
Alexandersson E, Mulugeta T, Lankinen Å, et al. (2016) Plant resistance inducers against pathogens in Solanaceae species-From molecular mechanisms to field application. Int J Mol Sci 17: 1673. doi: 10.3390/ijms17101673
|
| [91] | Borges AA, Sandalio LM (2015) Induced resistance for plant defense. Front Plant Sci 6: 109. |
| [92] |
Iriti M, Varoni EM (2017) Moving to the field: Plant innate immunity in crop protection. Int J Mol Sci 18: 640. doi: 10.3390/ijms18030640
|
| [93] |
Capanoglu E (2010) The potential of priming in food production. Trends Food Sci Technol 21: 399-407. doi: 10.1016/j.tifs.2010.05.001
|
| [94] | Aranega-Bou P, de la O Leyva M, Finiti I, et al. (2014) Priming of plant resistance by natural compounds. Hexanoic acid as a model. Front Plant Sci 5: 488. |
| [95] | Piquerez SJM, Harvey SE, Beynon JL, et al. (2014) Improving crop disease resistance: lessons from research on Arabidopsis and tomato. Front Plant Sci 5: 671. |
| [96] |
Van Aubel G, Cambier P, Dieu M, et al. (2016) Plant immunity induced by COS-OGA elicitor is a cumulative process that involves salicylic acid. Plant Sci 247: 60-70. doi: 10.1016/j.plantsci.2016.03.005
|
| [97] |
Garner CM, Kim SH, Spears BJ, et al. (2016) Express yourself: Transcriptional regulation of plant innate immunity. Sem Cell Dev Biol 56: 150-162. doi: 10.1016/j.semcdb.2016.05.002
|
| [98] |
Iriti M, Picchi V, Rossoni M (2009) Chitosan antitranspirant activity is due to abscisic acid-dependent stomatal closure. Environ Exp Bot 66: 493-500. doi: 10.1016/j.envexpbot.2009.01.004
|
| [99] |
Iriti M, Castorina G, Vitalini S, et al. (2010) Chitosan-induced ethylene-independent resistance does not reduce crop yield in bean. Biol Control 54: 241-247. doi: 10.1016/j.biocontrol.2010.05.012
|
| [100] |
Lamichhane JR, Bischoff-Schaefer M, Bluemel S, et al. (2017) Identifying obstacles and ranking common biological control research priorities for Europe to manage most economically important pests in arable, vegetable and perennial crops. Pest Manag Sci 73: 14-21. doi: 10.1002/ps.4423
|
| [101] | Gouveia BC, Calil IP, Machado JPB, et al. (2016) Immune receptors and co-receptors in antiviral innate immunity in plants. Front Microbiol 7: 2139. |
| [102] |
Brutus A, Sicilia F, Macone A, et al. (2010) A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides. Proc Natl Acad Sci USA 107: 9452-9457. doi: 10.1073/pnas.1000675107
|
| [103] | Cervone F, De Lorenzo G, Brutus A, et al. (2009) Constructs expressing chimeric receptors, and their use for controlled activation of defense responses to pathogens in plants. RM2009A000279 Patent. |
| [104] |
Rodriguez-Moreno L, Song Y, Thomma BPHJ (2017) Transfer and engineering of immune receptors to improve recognition capacities in plants. Curr Opin Plant Biol 38: 42-49. doi: 10.1016/j.pbi.2017.04.010
|
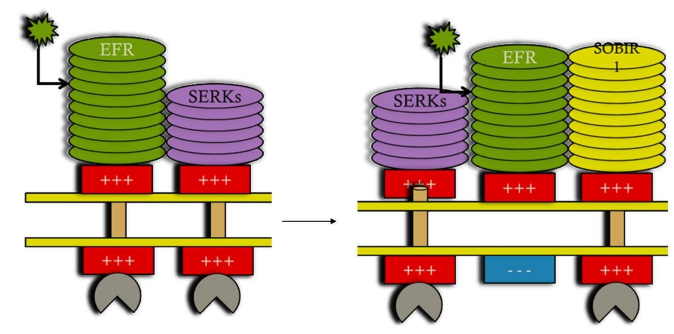
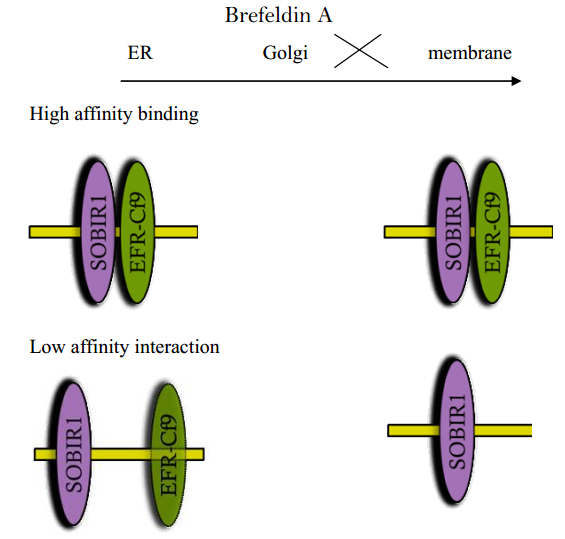
Figures(2)

Nataša Čerekovic, Palmiro Poltronieri. Plant signaling pathways activating defence response and interfering mechanisms by pathogen effectors, protein decoys and bodyguards[J]. AIMS Molecular Science, 2017, 4(3): 370-388. doi: 10.3934/molsci.2017.3.370







 DownLoad:
DownLoad: