Citation: M. Reza Jabalameli, Ignacio Briceno, Julio Martinez, Ignacio Briceno, Reuben J. Pengelly, Sarah Ennis, Andrew Collins. Aarskog-Scott syndrome: phenotypic and genetic heterogeneity[J]. AIMS Genetics, 2016, 3(1): 49-59. doi: 10.3934/genet.2016.1.49

| [1] | Aarskog D (1971) A familial syndrome of short stature associated with facial dysplasia and genital anomalies. Birth Defects Orig Artic Ser 7: 235-239. |
| [2] | Scott CI (1971) Unusual facies, joint hypermobility, genital anomaly and short stature: a new dysmorphic syndrome. Birth Defects Orig Artic Ser 7: 240-246. |
| [3] |
Orrico A, Galli L, Cavaliere ML, et al. (2004) Phenotypic and molecular characterisation of the Aarskog-Scott syndrome: a survey of the clinical variability in light of FGD1 mutation analysis in 46 patients. Eur J Hum Genet 12: 16-23. doi: 10.1038/sj.ejhg.5201081

|
| [4] |
Estrada L, Caron E, Gorski JL (2001) Fgd1, the Cdc42 guanine nucleotide exchange factor responsible for faciogenital dysplasia, is localized to the subcortical actin cytoskeleton and Golgi membrane. Hum Mol Genet 10: 485-495. doi: 10.1093/hmg/10.5.485
|
| [5] | Orrico A, Galli L, Clayton-Smith J, et al. (2011) Clinical utility gene card for: Aarskog-Scott syndrome (faciogenital dysplasia). Eur J Hum Genet 19. |
| [6] |
Orrico A, Galli L, Faivre L, et al. (2010) Aarskog-Scott Syndrome: Clinical Update and Report of Nine Novel Mutations of the FGD1 Gene. Am J Med Genet A 152a: 313-318. doi: 10.1002/ajmg.a.33199
|
| [7] |
Teebi AS, Rucquoi JK, Meyn MS (1993) Aarskog Syndrome - Report of a Family with Review and Discussion of Nosology. Am J Med Genet 46: 501-509. doi: 10.1002/ajmg.1320460508
|
| [8] |
Perez-Coria M, Lugo-Trampe JJ, Zamudio-Osuna M, et al. (2015) Identification of novel mutations in Mexican patients with Aarskog-Scott syndrome. Mol Genet Genomic Med 3: 197-202. doi: 10.1002/mgg3.132
|
| [9] |
Logie LJ, Porteous ME (1998) Intelligence and development in Aarskog syndrome. Arch Dis Child 79: 359-360. doi: 10.1136/adc.79.4.359
|
| [10] |
Orrico A, Galli L, Buoni S, et al. (2005) Attention-deficit/hyperactivity disorder (ADHD) and variable clinical expression of Aarskog-Scott syndrome due to a novel FGD1 gene mutation (R408Q). Am J Med Genet A 135a: 99-102. doi: 10.1002/ajmg.a.30700
|
| [11] | Nayak RB, Lambika, Bhogale GS, et al. (2012) Mania with Aarskog-Scott Syndrome. Indian Pediatrics 49: 327-328. |
| [12] |
Roberts AE, Allanson JE, Tartaglia M, et al. (2013) Noonan syndrome. Lancet 381: 333-342. doi: 10.1016/S0140-6736(12)61023-X
|
| [13] |
Al Kaissi A, Bieganski T, Baranska D, et al. (2007) Robinow syndrome: Report of two cases and review of the literature. Australas Radiol 51: 83-86. doi: 10.1111/j.1440-1673.2006.01668.x
|
| [14] |
Smpokou P, Zand DJ, Rosenbaum KN, et al. (2015) Malignancy in Noonan syndrome and related disorders. Clin Genet 88: 516-522. doi: 10.1111/cge.12568
|
| [15] |
Fryns JP (1992) Aarskog Syndrome - the Changing Phenotype with Age. Am J Med Genet 43: 420-427. doi: 10.1002/ajmg.1320430164
|
| [16] |
Galupa R, Heard E (2015) X-chromosome inactivation: new insights into cis and trans regulation. Curr Opin Genet Dev 31: 57-66. doi: 10.1016/j.gde.2015.04.002
|
| [17] |
Jogiya A, Sandy C (2005) Mild optic nerve hypoplasia with retinal venous tortuosity in aarskog (facial-digital-genital) syndrome. Ophthalmic Genet 26: 139-141. doi: 10.1080/13816810500229025
|
| [18] |
Andrassy RJ, Murthy S, Woolley MM (1979) Aarskog syndrome: significance for the surgeon. J Pediatr Surg 14: 462-464. doi: 10.1016/S0022-3468(79)80016-0
|
| [19] |
Mikelsaar RV, Lurie IW (1992) Atypical case of Aarskog syndrome. J Med Genet 29: 349-350. doi: 10.1136/jmg.29.5.349
|
| [20] | Fryns JP, Van den Berghe H (1989) On the occurrence of macroorchidism and mental handicap in the Aarskog syndrome. J Genet Hum 37: 221-223. |
| [21] |
Teebi AS, Naguib KK, Alawadi SA, et al. (1988) New Autosomal Recessive Faciodigitogenital Syndrome. J Med Genet 25: 400-406. doi: 10.1136/jmg.25.6.400
|
| [22] | Teebi AS, Alawadi SA (1991) Kuwait Type Faciodigitogenital Syndrome. J Med Genet 28: 805-805. |
| [23] |
Bawle E, Tyrkus M, Lipman S, et al. (1984) Aarskog Syndrome - Full Male and Female Expression Associated with an X-Autosome Translocation. Am J Med Genet 17: 595-602. doi: 10.1002/ajmg.1320170307
|
| [24] | Person AD, Beiraghi S, Sieben CM, et al. (2010) WNT5A mutations in patients with autosomal dominant Robinow syndrome. Dev Dyn 239: 327-337. |
| [25] |
Stevenson RE, May M, Arena JF, et al. (1994) Aarskog-Scott syndrome: confirmation of linkage to the pericentromeric region of the X chromosome. Am J Med Genet 52: 339-345. doi: 10.1002/ajmg.1320520317
|
| [26] |
Schmidt A, Hall A (2002) Guanine nucleotide exchange factors for Rho GTPases: turning on the switch. Genes Dev 16: 1587-1609. doi: 10.1101/gad.1003302
|
| [27] |
Zheng Y (2001) Dbl family guanine nucleotide exchange factors. Trends Biochem Sci 26: 724-732. doi: 10.1016/S0968-0004(01)01973-9
|
| [28] |
Genot E, Daubon T, Sorrentino V, et al. (2012) FGD1 as a central regulator of extracellular matrix remodelling - lessons from faciogenital dysplasia. J Cell Sci 125: 3265-3270. doi: 10.1242/jcs.093419
|
| [29] |
Grier RE, Farrington FH, Kendig R, et al. (1983) Autosomal dominant inheritance of the Aarskog syndrome. Am J Med Genet 15: 39-46. doi: 10.1002/ajmg.1320150105
|
| [30] |
Redin C, Le Gras S, Mhamdi O, et al. (2012) Targeted high-throughput sequencing for diagnosis of genetically heterogeneous diseases: efficient mutation detection in Bardet-Biedl and Alstrom syndromes. J Med Genet 49: 502-512. doi: 10.1136/jmedgenet-2012-100875
|
| [31] |
Alkuraya FS (2013) The application of next-generation sequencing in the autozygosity mapping of human recessive diseases. Hum Genet 132: 1197-1211. doi: 10.1007/s00439-013-1344-x
|
| [32] |
Pasteris NG, Cadle A, Logie LJ, et al. (1994) Isolation and Characterization of the Faciogenital Dysplasia (Aarskog-Scott Syndrome) Gene - a Putative Rho/Rac Guanine-Nucleotide Exchange Factor. Cell 79: 669-678. doi: 10.1016/0092-8674(94)90552-5
|
| [33] | Den Dunnen JT, LOVD v.3.0. 2015. Available from: http://www.lovd.nl/3.0/home |
| [34] |
Tartaglia M, Gelb BD, Zenker M (2011) Noonan syndrome and clinically related disorders. Best Pract Res Clin Endocrinol Metab 25: 161-179. doi: 10.1016/j.beem.2010.09.002
|
| [35] |
Aoki Y, Niihori T, Inoue S, et al. (2016) Recent advances in RASopathies. J Hum Genet 61: 33-39. doi: 10.1038/jhg.2015.114
|
| [36] |
Zou W, Greenblatt MB, Shim JH, et al. (2011) MLK3 regulates bone development downstream of the faciogenital dysplasia protein FGD1 in mice. J Clin Invest 121: 4383-4392. doi: 10.1172/JCI59041
|
| [37] |
Gao L, Gorski JL, Chen CS (2011) The Cdc42 guanine nucleotide exchange factor FGD1 regulates osteogenesis in human mesenchymal stem cells. Am J Pathol 178: 969-974. doi: 10.1016/j.ajpath.2010.11.051
|
| [38] |
von Mering C, Huynen M, Jaeggi D, et al. (2003) STRING: a database of predicted functional associations between proteins. Nucleic Acids Res 31: 258-261. doi: 10.1093/nar/gkg034
|
| [39] |
Aten E, Sun Y, Almomani R, et al. (2013) Exome sequencing identifies a branch point variant in Aarskog-Scott syndrome. Hum Mutat 34: 430-434. doi: 10.1002/humu.22252
|
| [40] |
Desmet FO, Hamroun D, Lalande M, et al. (2009) Human Splicing Finder: an online bioinformatics tool to predict splicing signals. Nucleic Acids Res 37: e67. doi: 10.1093/nar/gkp215
|
| [41] |
Taylor JC, Martin HC, Lise S, et al. (2015) Factors influencing success of clinical genome sequencing across a broad spectrum of disorders. Nat Genet 47: 717-726. doi: 10.1038/ng.3304
|
| [42] |
Rimmer A, Phan H, Mathieson I, et al. (2014) Integrating mapping-, assembly- and haplotype-based approaches for calling variants in clinical sequencing applications. Nat Genet 46: 912-918. doi: 10.1038/ng.3036
|
| [43] |
Pertz O (2010) Spatio-temporal Rho GTPase signaling - where are we now? J Cell Sci 123: 1841-1850. doi: 10.1242/jcs.064345
|
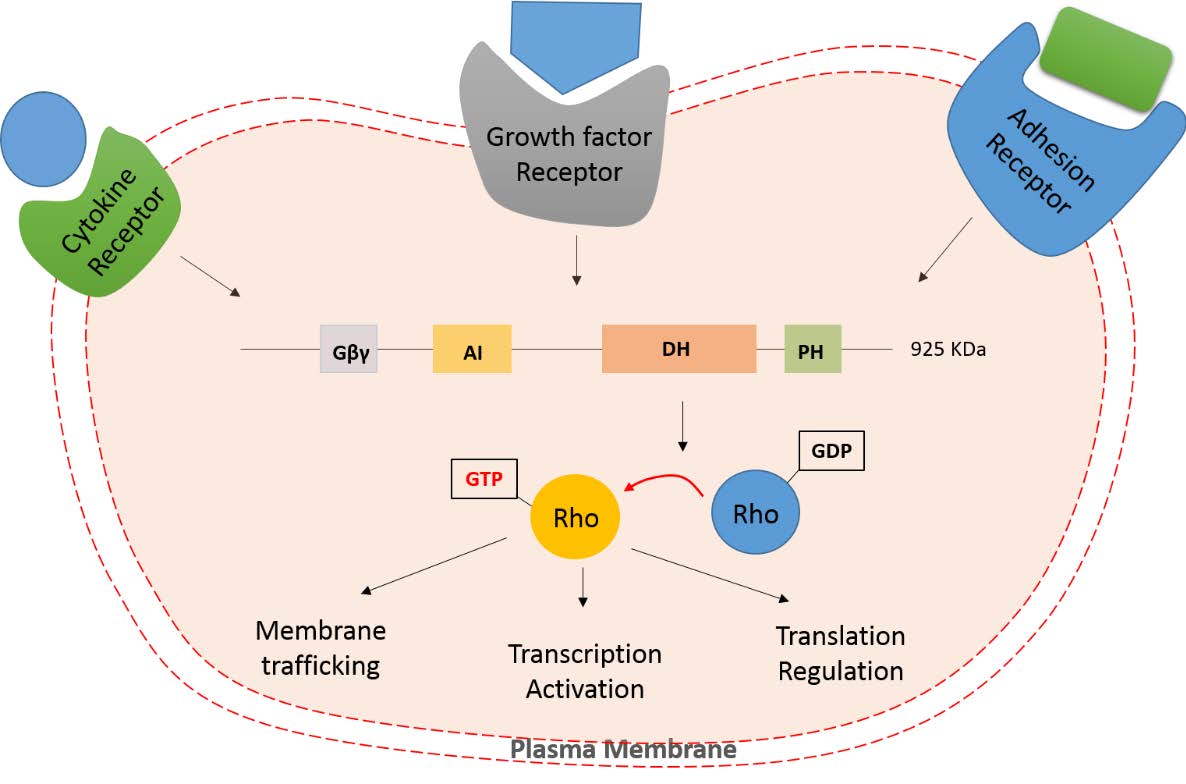
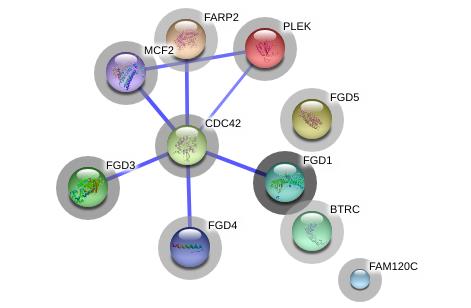
Figures(2) / Tables(2)

M. Reza Jabalameli, Ignacio Briceno, Julio Martinez, Ignacio Briceno, Reuben J. Pengelly, Sarah Ennis, Andrew Collins. Aarskog-Scott syndrome: phenotypic and genetic heterogeneity[J]. AIMS Genetics, 2016, 3(1): 49-59. doi: 10.3934/genet.2016.1.49







 DownLoad:
DownLoad: