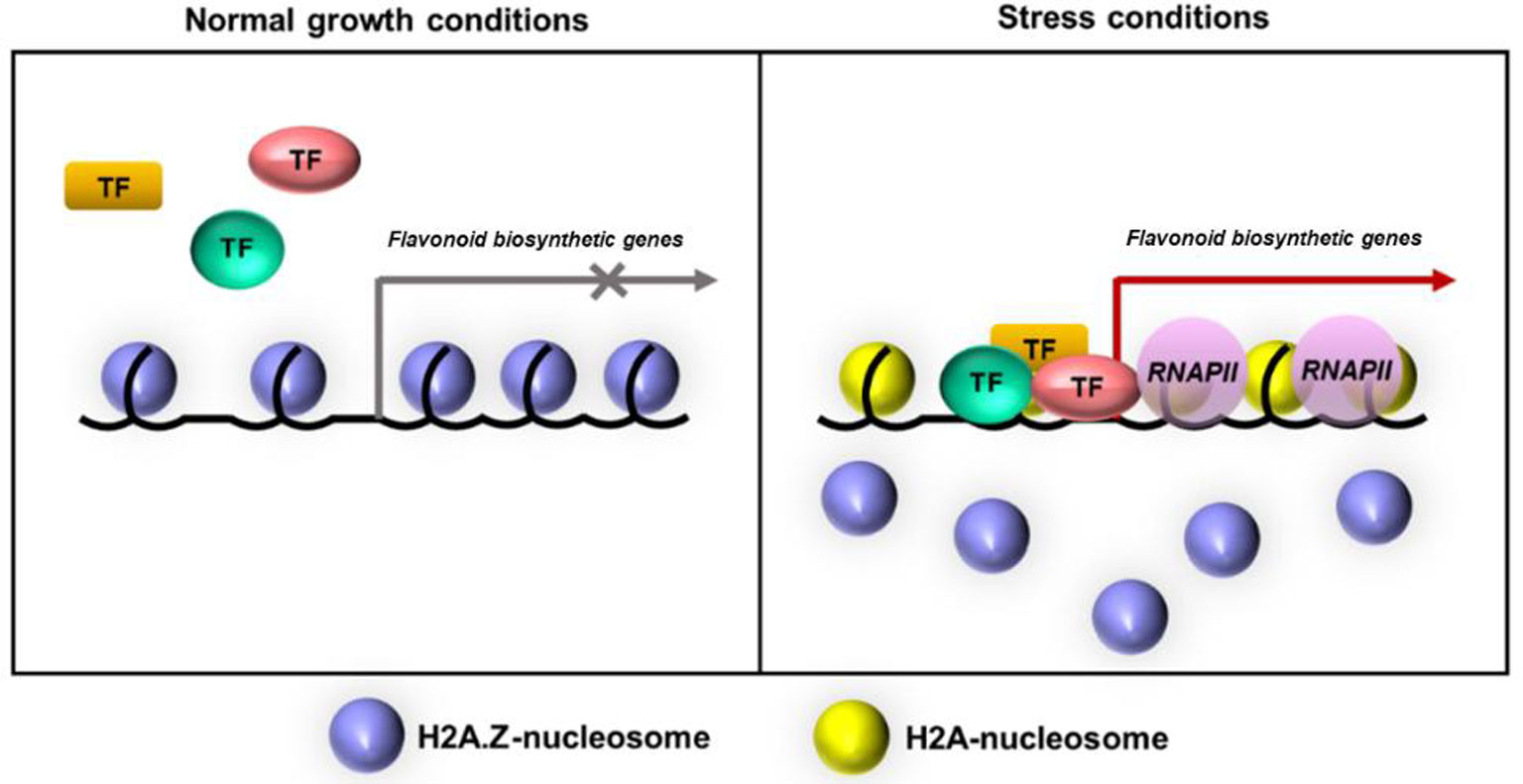
Flavonoid is an important group of plant secondary metabolites. The biosynthesis of this flavonoid group can be precisely regulated by many environmental factors. Currently, histone variants have been documented as important factors in the regulation of eukaryotic gene expression. H2A.Z histone variant has been found to function in numerous plant physiological programs including flavonoid biosynthesis. Moreover, the environmental changes have been shown to significantly influence the replacement between histone proteins and their variants leading to alterations in gene expression. Based on the recent studies, this mini-review is to provide an updated view on the functions of histone variant H2A.Z in the regulation of flavonoid biosynthetic gene expression. In addition, this also suggests a model in which the H2A.Z-containing nucleosomes can be evicted upon environmental stress conditions to facilitate the targeting of transcriptional activators to these flavonoid biosynthetic genes resulting in gene activation and flavonoid accumulation in Arabidopsis plants.
Citation: Nguyen Hoai Nguyen. Histone variant H2A.Z and transcriptional activators may antagonistically regulate flavonoid biosynthesis[J]. AIMS Bioengineering, 2020, 7(1): 55-59. doi: 10.3934/bioeng.2020005
Flavonoid is an important group of plant secondary metabolites. The biosynthesis of this flavonoid group can be precisely regulated by many environmental factors. Currently, histone variants have been documented as important factors in the regulation of eukaryotic gene expression. H2A.Z histone variant has been found to function in numerous plant physiological programs including flavonoid biosynthesis. Moreover, the environmental changes have been shown to significantly influence the replacement between histone proteins and their variants leading to alterations in gene expression. Based on the recent studies, this mini-review is to provide an updated view on the functions of histone variant H2A.Z in the regulation of flavonoid biosynthetic gene expression. In addition, this also suggests a model in which the H2A.Z-containing nucleosomes can be evicted upon environmental stress conditions to facilitate the targeting of transcriptional activators to these flavonoid biosynthetic genes resulting in gene activation and flavonoid accumulation in Arabidopsis plants.

| [1] |
Nakabayashi R, Yonekura-Sakakibara K, Urano K, et al. (2014) Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J 77: 367-379. doi: 10.1111/tpj.12388

|
| [2] |
Nguyen NH, Kim JH, Kwon J, et al. (2016) Characterization of Arabidopsis thaliana FLAVONOL SYNTHASE 1 (FLS1)-overexpression plants in response to abiotic stress. Plant Physiol Biochem 103: 133-142. doi: 10.1016/j.plaphy.2016.03.010
|
| [3] |
Peer WA, Murphy AS (2007) Flavonoids and auxin transport: modulators or regulators? Trends Plant Sci 12: 556-563. doi: 10.1016/j.tplants.2007.10.003
|
| [4] |
Winkel-Shirley B (2002) Biosynthesis of flavonoids and effects of stress. Curr Opin Plant Biol 5: 218-223. doi: 10.1016/S1369-5266(02)00256-X
|
| [5] |
Nguyen NH, Jeong CY, Kang GH, et al. (2015) MYBD employed by HY5 increases anthocyanin accumulation via repression of MYBL2 in Arabidopsis. Plant J 84: 1192-1205. doi: 10.1111/tpj.13077
|
| [6] |
Kim S, Hwang G, Lee S, et al. (2017) High ambient temperature represses anthocyanin biosynthesis through degradation of HY5. Front Plant Sci 8: 1787. doi: 10.3389/fpls.2017.01787
|
| [7] |
Cai HY, Zhang M, Chai MN, et al. (2019) Epigenetic regulation of anthocyanin biosynthesis by an antagonistic interaction between H2A.Z and H3K4me3. New Phytol 221: 295-308. doi: 10.1111/nph.15306
|
| [8] |
Mehrtens F, Kranz H, Bednarek P, et al. (2005) The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol 138: 1083-1096. doi: 10.1104/pp.104.058032
|
| [9] |
Stracke R, Ishihara H, Huep G, et al. (2007) Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J 50: 660-677. doi: 10.1111/j.1365-313X.2007.03078.x
|
| [10] |
Richmond TJ, Finch JT, Rushton B, et al. (1984) Structure of the nucleosome core particle at 7Å resolution. Nature 311: 532-537. doi: 10.1038/311532a0
|
| [11] | Kim JM, Sasaki T, Ueda M, et al. (2015) Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front Plant Sci 6: 114. |
| [12] |
Lai WKM, Pugh BF (2017) Understanding nucleosome dynamics and their links to gene expression and DNA replication. Nat Rev Mol Cell Bio 18: 548-562. doi: 10.1038/nrm.2017.47
|
| [13] |
Kumar SV, Wigge PA (2010) H2A.Z-containing nucleosomes mediate the thermosensory response in Arabidopsis. Cell 140: 136-147. doi: 10.1016/j.cell.2009.11.006
|
| [14] |
Smith AP, Jain A, Deal RB, et al. (2010) Histone H2A.Z regulates the expression of several classes of phosphate starvation response genes but not as a transcriptional activator. Plant Physiol 152: 217-225. doi: 10.1104/pp.109.145532
|
| [15] |
Sura W, Kabza M, Karlowski WM, et al. (2017) Dual role of the histone variant H2A. Z in transcriptional regulation of stress-response genes. Plant Cell 29: 791-807. doi: 10.1105/tpc.16.00573
|
| [16] |
Nguyen NH, Cheong JJ (2018) H2A.Z-containing nucleosomes are evicted to activate AtMYB44 transcription in response to salt stress. Biochem Biophys Res Commun 499: 1039-1043. doi: 10.1016/j.bbrc.2018.04.048
|
| [17] |
Hu GQ, Cui KR, Northrup D, et al. (2013) H2A. Z facilitates access of active and repressive complexes to chromatin in embryonic stem cell self-renewal and differentiation. Cell Stem Cell 12: 180-192. doi: 10.1016/j.stem.2012.11.003
|
| [18] |
Schuettengruber B, Chourrout D, Vervoort M, et al. (2007) Genome regulation by polycomb and trithorax proteins. Cell 128: 735-745. doi: 10.1016/j.cell.2007.02.009
|
| [19] |
Xu ML, Leichty AR, Hu TQ, et al. (2018) H2A.Z promotes the transcription of MIR156A and MIR156C in Arabidopsis by facilitating the deposition of H3K4me3. Development 145: dev152868. doi: 10.1242/dev.152868
|
| [20] |
Dai X, Bai Y, Zhao L, et al. (2017) H2A. Z represses gene expression by modulating promoter nucleosome structure and enhancer histone modifications in Arabidopsis. Mol Plant 10: 1274-1292. doi: 10.1016/j.molp.2017.09.007
|
| [21] |
Kumar SV, Lucyshyn D, Jaeger KE, et al. (2012) Transcription factor PIF4 controls the thermosensory activation of flowering. Nature 484: 242-245. doi: 10.1038/nature10928
|
| [22] |
Borevitz JO, Xia YJ, Blount J, et al. (2000) Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant Cell 12: 2383-2393. doi: 10.1105/tpc.12.12.2383
|
| [23] |
Shin J, Park E, Choi G (2007) PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. Plant J 49: 981-994. doi: 10.1111/j.1365-313X.2006.03021.x
|
Figures(1)

Nguyen Hoai Nguyen. Histone variant H2A.Z and transcriptional activators may antagonistically regulate flavonoid biosynthesis[J]. AIMS Bioengineering, 2020, 7(1): 55-59. doi: 10.3934/bioeng.2020005







 DownLoad:
DownLoad: