Citation: Nieves Barros, Sergio Feijoo, César Pérez-Cruzado, Lee D. Hansen. Effect of soil storage at 4 °C on the calorespirometric measurements of soil microbial metabolism[J]. AIMS Microbiology, 2017, 3(4): 762-773. doi: 10.3934/microbiol.2017.4.762

| [1] | Šimek M, Santruckova H (1999) Influence of storage of soil samples on microbial biomass and its activity. Rost Vyroba 44: 415–419. |
| [2] |
Yanai Y, Toyota K, Okazaki M (2007) Response of denitrifying communities to successive soil freeze-thaw cycles. Biol Fert Soils 44: 113–119. doi: 10.1007/s00374-007-0185-y

|
| [3] |
Lauber CL, Zhou N, Gordon JI, et al. (2010) Effect of storage conditions on the assessment of bacterial community structure in soil and human-associated samples. FEMS Microbiol Lett 307: 80–86. doi: 10.1111/j.1574-6968.2010.01965.x
|
| [4] | Lee YB, Lorenz N, Dick LK, et al. (2006) Cold storage and pre-treatment incubation effects on soil microbial properties. Soil Sci Soc Am J 71: 1299–1305. |
| [5] | Rubin BER, Gibbons SM, Kennedy S, et al. (2013) Investigating the impact of storage conditions on microbial community composition in soil samples. PLoS One 8: 1–9. |
| [6] |
Stenberg B, Johansson M, Pell M, et al. (1998) Microbial biomass and activities in soils as affected by frozen and cold storage. Soil Biol Biochem 30: 393–402. doi: 10.1016/S0038-0717(97)00125-9
|
| [7] |
Larsen KS, Jonasson S, Michelsen A (2002) Repeated freeze-thaw cycles and their effects on biological processes in two arctic ecosystem types. Appl Soil Ecol 21: 187–195. doi: 10.1016/S0929-1393(02)00093-8
|
| [8] |
Barros N, Salgado J, Feijoo S (2007) Calorimetry and soil. Thermochim Acta 458: 11–17. doi: 10.1016/j.tca.2007.01.010
|
| [9] |
Rong XM, Huang QY, Jiang DH, et al. (2007) Isothermal microcalorimetry: A review of application in soil and environmental sciences. Pedosphere 17: 137–145. doi: 10.1016/S1002-0160(07)60019-8
|
| [10] |
Xu J, Feng Y, Barros N, et al. (2017) Exploring the potential of microcalorimetry to study soil microbial metabolic diversity. J Therm Anal Calorim 127: 1457–1465. doi: 10.1007/s10973-016-5952-2
|
| [11] |
Barros N, Hansen LD, Piñeiro V, et al. (2016) Calorimetry measures the response of soil organic matter biodegradation to increasing temperature. J Therm Anal Calorim 123: 2397–2403. doi: 10.1007/s10973-015-4947-8
|
| [12] | Barros N, Feijoo S, Balsa R, et al. (2017) Calorimetry reveals the response of soil microbial metabolism to increasing temperature in soils with different thermal, chemical and biological properties. Adv Mat Tech Env 1: 27–37. |
| [13] |
Barros N, Feijoo S (2003) A combined mass and energy balance to provide bioindicators of soil microbiological quality. Biophys Chem 104: 561–572. doi: 10.1016/S0301-4622(03)00059-0
|
| [14] |
Harris JA, Ritz K, Coucheney E, et al. (2012) The thermodynamic efficiency of soil microbial communities subject to long-term stress is lower than those under conventional input regimes. Soil Biol Biochem 47: 149–157. doi: 10.1016/j.soilbio.2011.12.017
|
| [15] |
Wadsö L, Hansen LD (2015) Calorespirometry of terrestrial organisms and ecosystems. Methods 76: 11–19. doi: 10.1016/j.ymeth.2014.10.024
|
| [16] |
Barros N, Feijoo S, Hansen L (2011) Calorimetric determination of metabolic heat, CO2 rates and the calorespirometric ratio of soil basal metabolism. Geoderma 160: 542–547. doi: 10.1016/j.geoderma.2010.11.002
|
| [17] |
Herrmann AM, Bölscher T (2015) Simultaneous screening of microbial energetics and CO2 respiration in soil samples from different ecosystems. Soil Biol Biochem 83: 88–92. doi: 10.1016/j.soilbio.2015.01.020
|
| [18] |
Brueckner D, Solokhina A, Krähenbühl S, et al. (2017) A combined application of tunable diode laser absorption spectroscopy and isotherm micro-calorimetry for calorespirometric analysis. J Microbiol Meth 139: 210. doi: 10.1016/j.mimet.2017.06.012
|
| [19] |
Barros N, Hansen LD, Piñeiro V, et al. (2016) Factors influencing the calorespirometric ratios of soil microbial metabolism. Soil Biol Biochem 92: 221–229. doi: 10.1016/j.soilbio.2015.10.007
|
| [20] |
Barros N, Piñeiro V, Hansen LD (2015) Calorespirometry: A novel tool to assess the effect of temperature on soil organic matter decomposition. Thermochim Acta 618: 15–17. doi: 10.1016/j.tca.2015.09.005
|
| [21] |
Hansen LD, Macfarlane C, McKinnon N, et al. (2004) Use of calorespirometric ratios, heat per CO2 and heat per O2, to quantify metabolic paths and energetics of growing cells.Thermochim Acta 422: 55–61. doi: 10.1016/j.tca.2004.05.033
|
| [22] | Barros N, Merino A, Martín-Pastor M, et al. (2014) Changes in soil organic matter in a forestry chronosequence monitored by thermal analysis and calorimetry. SJSS 4: 239–253. |
| [23] |
Núñez-Regueira L, Barros N, Barja I (1994) Effect of storage of soil at 4 °C on the microbial activity studied by microcalorimetry. J Therm Anal Calorim 41: 1379–1383. doi: 10.1007/BF02549932
|
| [24] |
Pérez-Cruzado C, Sande B, Omil B, et al. (2014) Organic matter properties in soils afforested with Pinus radiata. Plant Soil 374: 381–398. doi: 10.1007/s11104-013-1896-5
|
| [25] |
Plante AF, Fernández JM, Haddix ML, et al. (2011) Biological, chemical and thermal indices of soil organic matter stability in four grasslands soils. Soil Biol Biochem 43: 1051–1058. doi: 10.1016/j.soilbio.2011.01.024
|
| [26] |
Pesaro M, Nicollier G, Zeyer J, et al. (2004) Impact of soil drying-rewetting stress on microbial communities and activities and on degradation of two crop protection products. Appl Environ Microb 70: 2577–2587. doi: 10.1128/AEM.70.5.2577-2587.2004
|
| [27] | R Core Team, R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria, 2015. Available from: https://www.R-project.org/. |
| [28] |
Lebuhn M, Heilmann B, Hartmann A (1994) Effects of drying/rewetting stress on microbial auxin production and L-tryptophan catabolism. Biol Fert Soils 18: 302–310. doi: 10.1007/BF00570633
|
| [29] |
Coxson ES, Parkinson D (1987) Winter respiratory activity in aspen woodland forest floor litter and soils. Soil Biol Biochem 19: 49–59. doi: 10.1016/0038-0717(87)90125-8
|
| [30] |
Nunan L, Lerch TZ, Pouteau V, et al. (2015) Metabolising old soil carbon: simply a matter of simply organic matter? Soil Biol Biochem 88: 128–136. doi: 10.1016/j.soilbio.2015.05.018
|
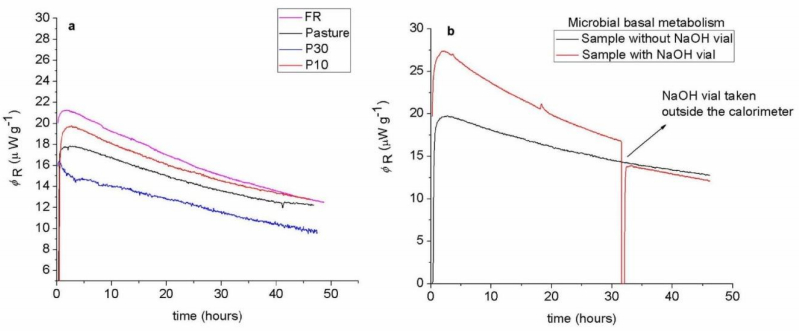
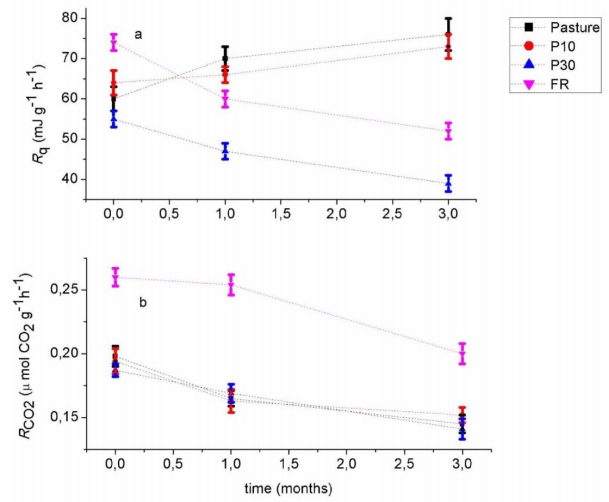
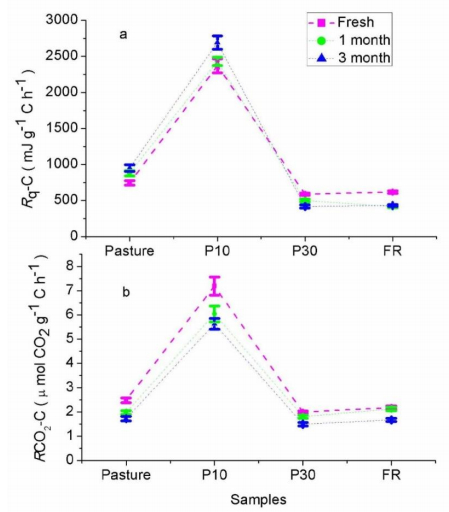
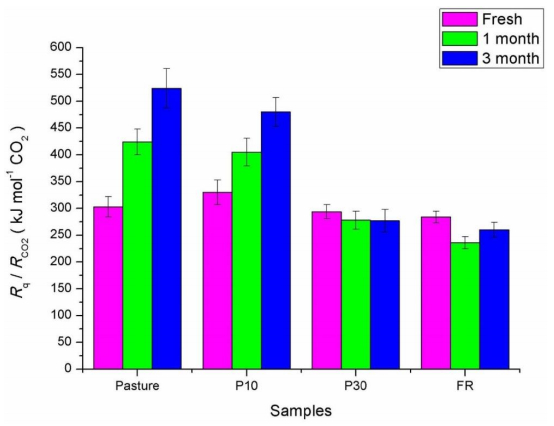
Figures(4) / Tables(1)

Nieves Barros, Sergio Feijoo, César Pérez-Cruzado, Lee D. Hansen. Effect of soil storage at 4 °C on the calorespirometric measurements of soil microbial metabolism[J]. AIMS Microbiology, 2017, 3(4): 762-773. doi: 10.3934/microbiol.2017.4.762







 DownLoad:
DownLoad: