Citation: Ioannis Panagopoulos, Athanassios Karayannis, Georgios Gouvalias, Nikolaos Karayannis, Pavlos Kassomenos. Chromium and nickel in the soils of industrial areas at Asopos river basin[J]. AIMS Environmental Science, 2016, 3(3): 420-438. doi: 10.3934/environsci.2016.3.420

| [1] |
Economou-Eliopoulos M, Megremi I, Vasilatos C (2011) Factors controlling the heterogeneous distribution of Cr(VI) in soil, plants and groundwater: evidence from the Asopos basin,Greece, Chem. Erde Geochem 71: 39-52. doi: 10.1016/j.chemer.2011.01.001

|
| [2] | Papanikolaou D (2009) Timing of tectonic emplacement of the Ophiolites and Terrane Paleogeography in the Hellenides, Lithos 108: 262-280. |
| [3] |
Moraetis D, Nikolaidis NP, Karatzas GP, et al. (2012) Origin and mobility of hexavalent chromium in North-Eastern Attica, Greece. Appl Geochem 27: 1170-1178. doi: 10.1016/j.apgeochem.2012.03.005
|
| [4] |
Valeton I, Biermann M, Reche R, et al. (1987) Genesis of Ni-laterites and bauxites inGreeceduring the Jurassic and Cretaceous, and their relation to ultrabasic parent rocks. ORE Geol Rev 2: 359-404. doi: 10.1016/0169-1368(87)90011-4
|
| [5] |
Eliopoulos D, Economou-Eliopoulos M (2000) Geochemical and mineralogical characteristics of Fe-Ni and bauxitic-laterite deposits ofGreece. Ore Geol Rev 16: 41-58. doi: 10.1016/S0169-1368(00)00003-2
|
| [6] | Oze C, Fendorf SE, Bird DK, et al. (2004) Chromium geochemistry of serpentine ultramafic rocks and serpentine soils from the Franciscan complex of California. Am J Sci 304: 67-101. |
| [7] |
Morrison JM, Goldhaber MB, Lee L, et al. (2009) A regional-scale study of chromium and nickel in soils of northern California, USA. Appl Geochem 24: 1500-1511. doi: 10.1016/j.apgeochem.2009.04.027
|
| [8] |
JBall JW, Izbicki JA (2004) Occurence of hexavalent chromium in ground water in the western Mojave desert, California. Appl Geochem 19: 1123-1135. doi: 10.1016/j.apgeochem.2004.01.011
|
| [9] | Becquer T, Quantin C, Sicot M, et al. (2003) Chromium availability in ultramafic soils from New Caledonia. Sci Total Environ 301: 251-261. |
| [10] | Cooper GRC (2000) Oxidation and toxicity of chromium in ultramafic soils inZimbabwe. Appl Geochem 17: 981-986. |
| [11] | Fantoni D, Brozzo G, Canepa M, et al. (2002) Natural hexavalent chromium in groundwater interacting with ophiolitic rocks. Environ Geol 42: 871-882. |
| [12] | Botsou F, Karageorgis AP, Dassenakis E, et al. (2011) Assessment of heavy metal contamination and mineral magnetic characterization of the Asopos River sediments (Central Greece), Mar Pollut Bull 62: 547-563. |
| [13] | Sybilla Ltd (2009) Soil investigation study, HAIS.A.(in Greek). Technical Soil Pollution Investigation Report submitted to and approved by the Environmental Directorate of the Region of Sterea Ellada. |
| [14] | Sybilla Ltd (2011) Soil investigation study, Europa Profile Aluminium S.A.(in Greek). Technical Soil Pollution Investigation Report submitted to and approved by the Environmental Directorate of the Region of Sterea Ellada. |
| [15] | Sybilla Ltd (2012) Soil investigation study, AlumincoS.A.(in Greek). Technical Soil Pollution Investigation Report submitted to and approved by the Environmental Directorate of the Region of Sterea Ellada. |
| [16] | Sybilla Ltd (2012), Soil investigation study, ViometaleS.A.(in Greek). Technical Soil Pollution Investigation Report submitted to and approved by the Environmental Directorate of the Region of Sterea Ellada. |
| [17] | LIFE10 ENV/GR/000601. CHARM, "Chromium in Asopos groundwater system: remediation technologies and measures", Technical Report, Deliverable 4.2: Results of supplementary monitoring programme in the groundwater system of Asopos, 2012. Available from: http://www.charm-life.gr/charm/index.php/en/documents. |
| [18] | Directive 2010/75/EU of the European Parliament and the Council on industrial emissions (the Industrial Emissions Directive or IED). |
| [19] | Sybilla Ltd (2015) Baseline Site Soil and Groundwater Pollution investigation studies, Europa Profile Aluminium S.A. (in Greek). Report submitted to and approved by the Environmental Directorate of the Region of Sterea Ellada |
| [20] | Sybilla Ltd (2015) Baseline Site Soil and Groundwater Pollution investigation studies, Aluminco S.A. (in Greek). Report submitted to and approved by the Environmental Directorate of the Region of Sterea Ellada |
| [21] | Panagopoulos I, Karayannis A, Kollias K, et al. (2015) Investigation of potential soil contamination with Cr and Ni in four metal finishing facilities at Asopos industrial area, J Hazard Mater 581, 20-26. |
| [22] | Salminen R (2005) Geochemical Atlas of Europe, ed., FOREGS, Geological Survey ofFinland, 2005. |
| [23] | Antibachi D, Kelepertzis E, Kelepertsis A (2012) Heavy Metals in Agricultural Soils of the Mouriki-Thiva Area (Central Greece) and Environmental Impact Implications. Soil Sediment Contam 21: 434-450. |
| [24] | Kanellopoulos C, Argyraki A, Mitropoulos P (2015) Geochemistry of serpentine agricultural soil and associated groundwater chemistry and vegetation in the area of Atalanti,Greece. J Geochem Explor 158: 22-33. |
| [25] | Carlon C, Derivation methods of soil screening values in Europe. A review and evaluation of national procedures towards harmonisation, ed. EC JRC EUR 22805-EN, Ispra, 2007. |
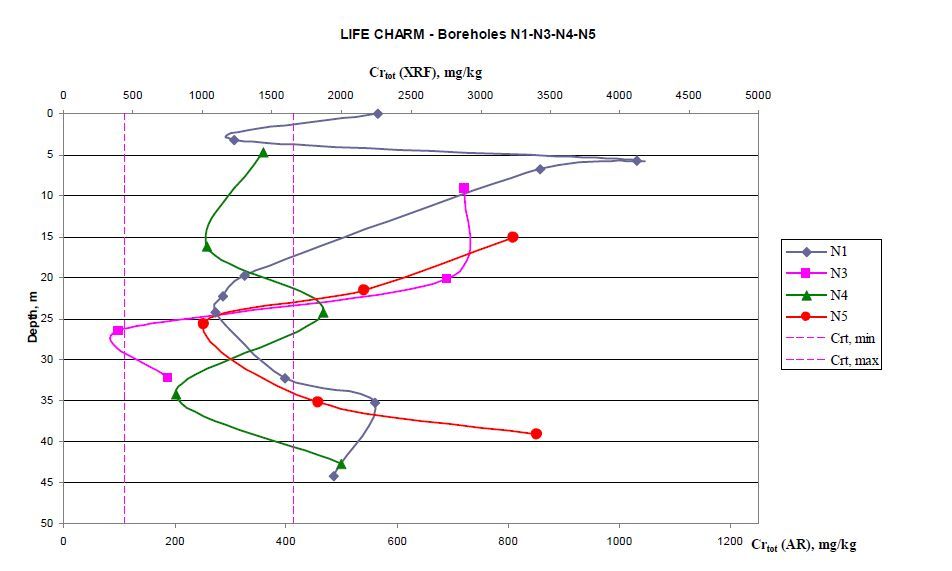
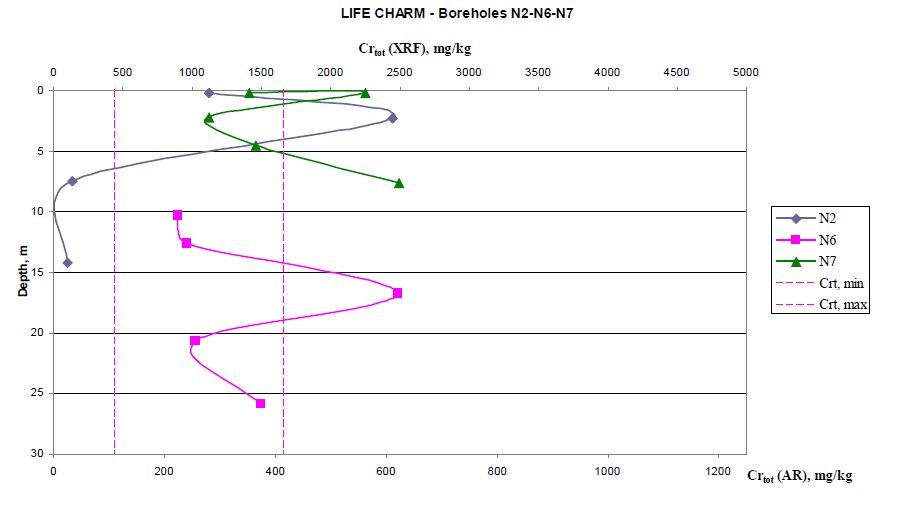
Figures(10) / Tables(6)

Ioannis Panagopoulos, Athanassios Karayannis, Georgios Gouvalias, Nikolaos Karayannis, Pavlos Kassomenos. Chromium and nickel in the soils of industrial areas at Asopos river basin[J]. AIMS Environmental Science, 2016, 3(3): 420-438. doi: 10.3934/environsci.2016.3.420







 DownLoad:
DownLoad: