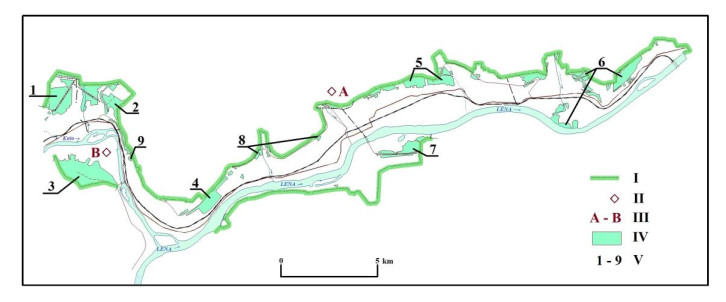
The Ust-Kutsky district and its center, the Ust-Kut city, have become the fastest growing in the Irkutsk region of Russia in the last decade. Ust-Kut is a very important settlement for the economic development of the northern territories of the Irkutsk region and the southern territories of the Republic of Sakha (Yakutia). The city is a key transport hub, where various types of transport routes converge: railway, aviation, automobile, river, and product pipeline. There are deposits of various minerals near Ust-Kut. The timber mining and timber-processing industries are represented here as much as possible. On the other hand, Ust-Kut is one of the ten most air-polluted cities in the Irkutsk region. Most of the air pollution consists of substances formed as a result of fuel combustion. The unfavorable air regime of the city is influenced by climatic conditions: relatively weak winds and temperature inversions in winter. The oil and gas deposits located nearby may help solve the problem of air pollution by gasification of Ust-Kut. In addition, water and soil pollution have been noted in the city due to technological problems of wastewater disposal, garbage collection, and recycling, as well as certain types of industrial waste processing. Therefore, the main task of the sustainable development of the city is to solve its environmental problems. Ust-Kut's environmental problems are solved at the expense of financial resources from government programs, which are not sufficient. But the tax on mining is completely federal, as these funds do not remain in the territory. Through this research, we have searched for ways to achieve a balance between economic development and environmental well-being in Ust-Kut. Specifically, we have explored additional types of economic activities that are safe for nature and public health. For Ust-Kut, located in the north of Eastern Siberia, it is important to increase the area of green spaces within the central part of the city. The development of a system of parks and squares involves their improvement and the formation of a connection into a single natural framework through linear landscaping objects and connections with urban forest sites. The use of sanitary protection zones around enterprises that emit into the environment will contribute to the formation of a comfortable environment and expand landscaping opportunities.
Citation: Tatyana P. Kalikhman. The economic situation and ways to solve the environmental problems in the city of Ust-Kut[J]. Urban Resilience and Sustainability, 2024, 2(2): 203-214. doi: 10.3934/urs.2024010
The Ust-Kutsky district and its center, the Ust-Kut city, have become the fastest growing in the Irkutsk region of Russia in the last decade. Ust-Kut is a very important settlement for the economic development of the northern territories of the Irkutsk region and the southern territories of the Republic of Sakha (Yakutia). The city is a key transport hub, where various types of transport routes converge: railway, aviation, automobile, river, and product pipeline. There are deposits of various minerals near Ust-Kut. The timber mining and timber-processing industries are represented here as much as possible. On the other hand, Ust-Kut is one of the ten most air-polluted cities in the Irkutsk region. Most of the air pollution consists of substances formed as a result of fuel combustion. The unfavorable air regime of the city is influenced by climatic conditions: relatively weak winds and temperature inversions in winter. The oil and gas deposits located nearby may help solve the problem of air pollution by gasification of Ust-Kut. In addition, water and soil pollution have been noted in the city due to technological problems of wastewater disposal, garbage collection, and recycling, as well as certain types of industrial waste processing. Therefore, the main task of the sustainable development of the city is to solve its environmental problems. Ust-Kut's environmental problems are solved at the expense of financial resources from government programs, which are not sufficient. But the tax on mining is completely federal, as these funds do not remain in the territory. Through this research, we have searched for ways to achieve a balance between economic development and environmental well-being in Ust-Kut. Specifically, we have explored additional types of economic activities that are safe for nature and public health. For Ust-Kut, located in the north of Eastern Siberia, it is important to increase the area of green spaces within the central part of the city. The development of a system of parks and squares involves their improvement and the formation of a connection into a single natural framework through linear landscaping objects and connections with urban forest sites. The use of sanitary protection zones around enterprises that emit into the environment will contribute to the formation of a comfortable environment and expand landscaping opportunities.

| [1] |
Dzhamalov RG, Reshetnyak OS, Vlasov KG, et al. (2021) Specific features of water chemistry and quality in the Lena River basin. Water Ecol 26: 33–43. http://dx.doi.org/10.23968/2305-3488.2021.26.3.33-43 doi: 10.23968/2305-3488.2021.26.3.33-43

|
| [2] | Kolesnikov SI, Kazeev KS, Tatlok RK, et al. (2014) Biodiagnostics of brown forest soils' resistance to oil pollution and heavy metals pollution in Western Caucasus. Siberian Ecol J 21: 493–500. Available from: https://innoscope.ru/analytics/publications/4168/?ysclid = lyr79tb4px46644963. |
| [3] |
Buluktaev AA (2017) Phytotoxicity and enzymatic activity in soils of Kalmykia under the influence of oil pollution. South Russ: Ecol Dev 12: 147–156. https://doi.org/10.18470/1992-1098-2017-4-147-156 doi: 10.18470/1992-1098-2017-4-147-156
|
| [4] | Kislitsina MN, Borisova GG (2016) Responses of Lemna minor L. to the exogenous phenolic compounds action. Scientific Notes of Petrozavodsk State University 157: 54–58. Available from: https://cyberleninka.ru/article/n/otvetnye-reaktsii-lemna-minor-l-na-deystvie-ekzogennyh-fenolnyh-soedineniy. |
| [5] | Borlakova TM, Semenov DS (2020) The role of the mineral extraction tax in the Russian budget system. Vector Econ. Available from: http://www.vectoreconomy.ru/images/publications/2020/9/taxes/Borlakova_Semenov.pdf. |
| [6] | Belovezhets LA, Volchatova IV, Medvedeva SA (2010) Promising methods of processing secondary lignocellulose raw materials. Chem Plant Raw Mater 5–16. Available from: https://cyberleninka.ru/article/n/perspektivnye-sposoby-pererabotki-vtorichnogo-lignotsellyuloznogo-syrya/viewer. |
| [7] |
Gao D, Zhou X, Mo X, et al. (2024) Unlocking sustainable growth: exploring the catalytic role of green finance in firms' green total factor productivity. Environ Sci Pollut Res 31: 14762–14774. https://doi.org/10.1007/s11356-024-32106-6 doi: 10.1007/s11356-024-32106-6
|
| [8] | Code of Rules "Territories improvement". Approved by Order of the Ministry of Construction and Housing and Communal Services of the Russian Federation No. 972-pr dated 12/16/2016. Available from: https://docs.cntd.ru/document/456054208. |
| [9] | Code of Rules "Building codes and regulations 2.07. 01-89 Urban development. Urban and rural planning and development" (2016b). Approved by Order of the Ministry of Construction and Housing and Communal Services of the Russian Federation No. 10/34-pr dated 12/30/2016. Available from: https://docs.cntd.ru/document/456054209. |
| [10] | Kalikhman TP, Bogdanov VN, Ogorodnikova LYu (2012) Specially Protected Natural Areas of the Siberian Federal District, Irkutsk (Russia): Ottisk Publishing House, 384. Available from: http://savesteppe.org/ru/archives/10541. |

Figures(2) / Tables(2)
Tatyana P. Kalikhman. The economic situation and ways to solve the environmental problems in the city of Ust-Kut[J]. Urban Resilience and Sustainability, 2024, 2(2): 203-214. doi: 10.3934/urs.2024010







 DownLoad:
DownLoad: