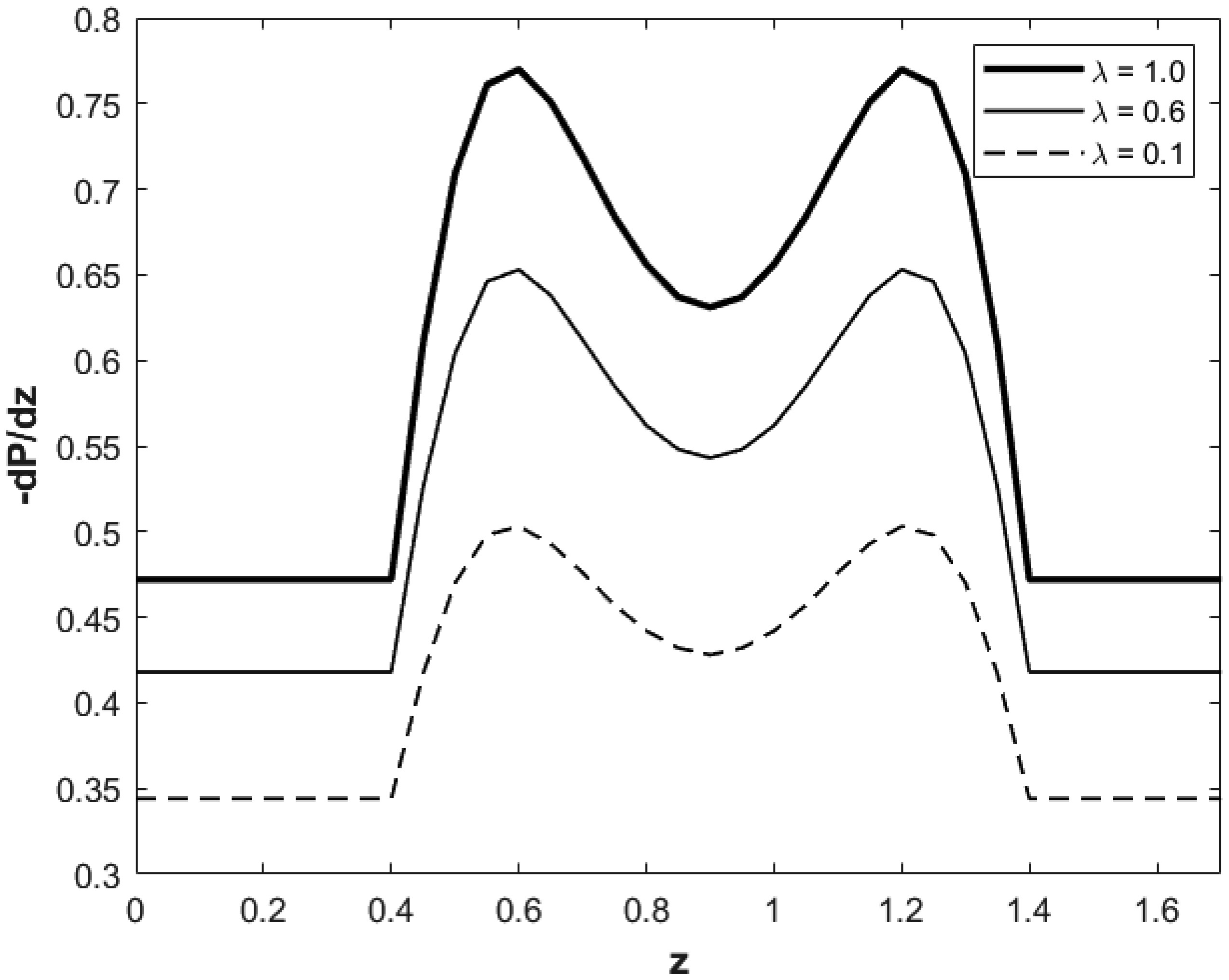
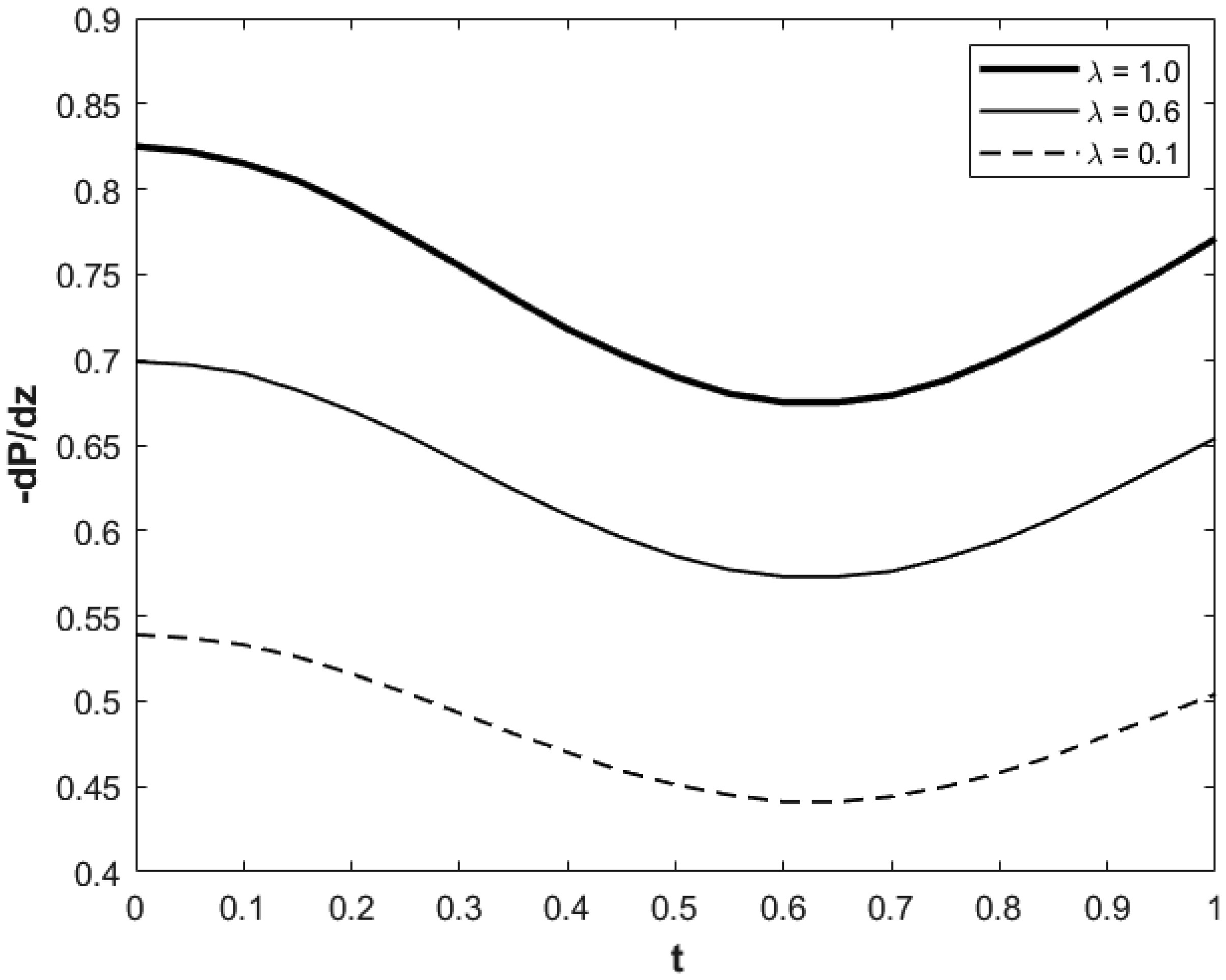
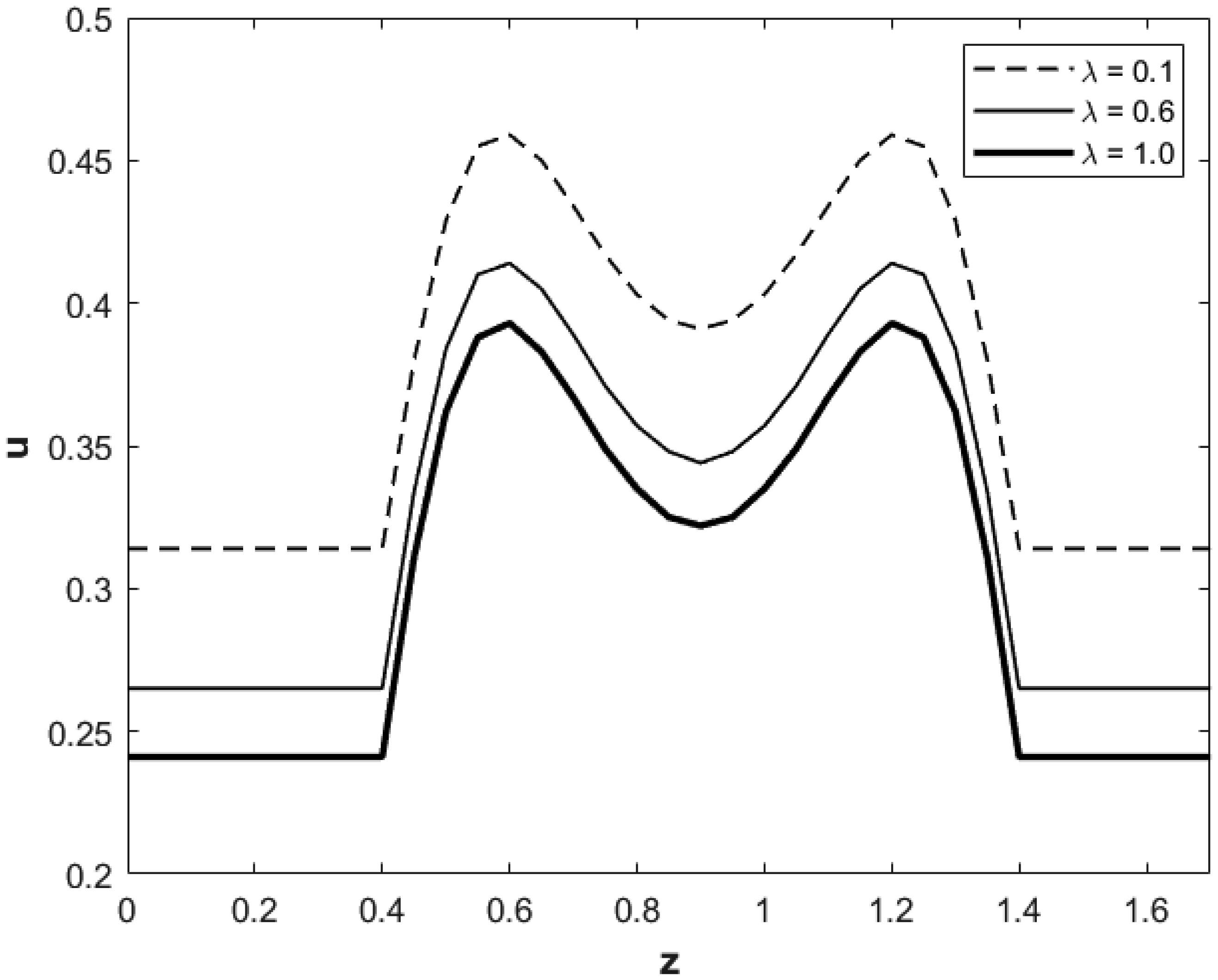
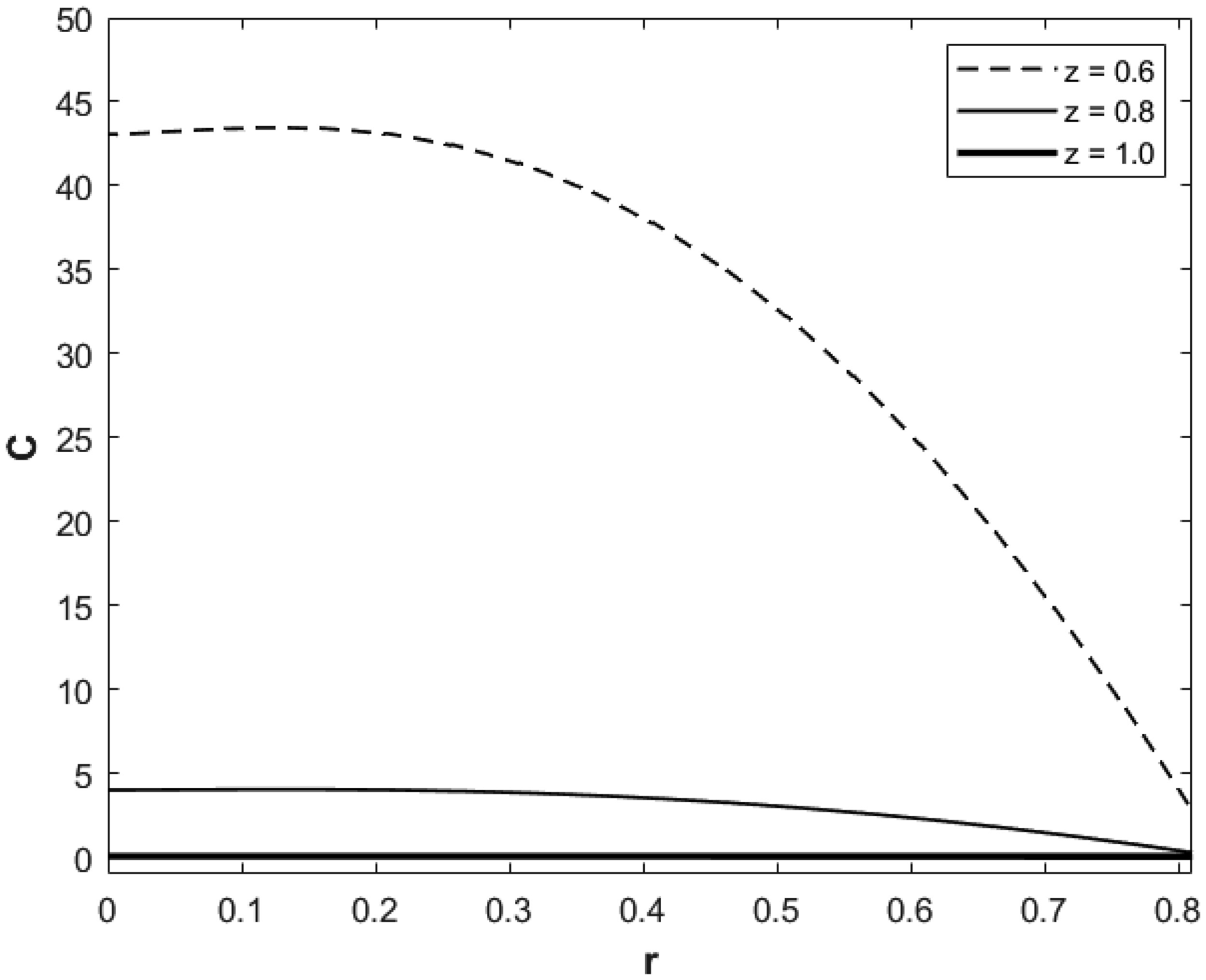
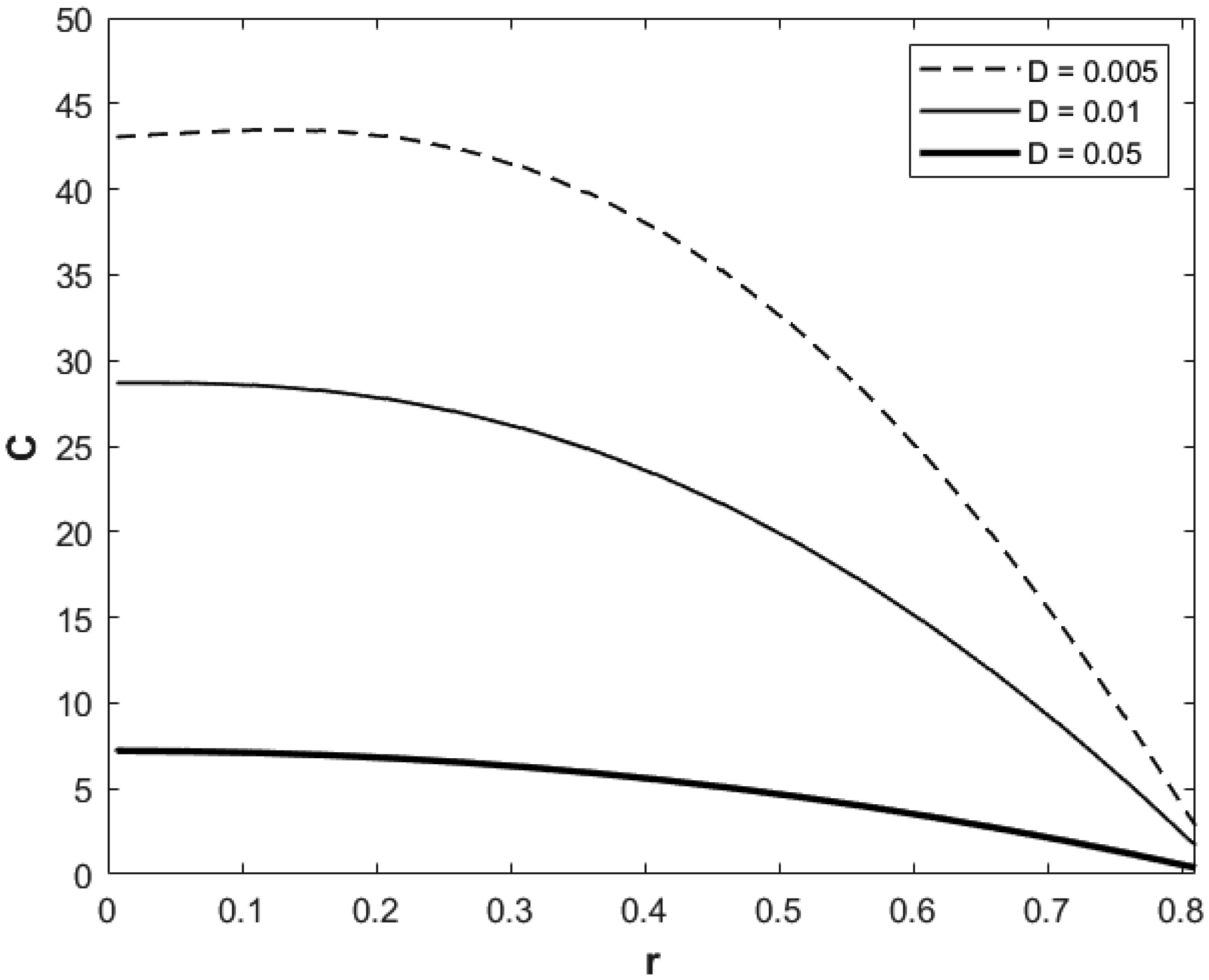
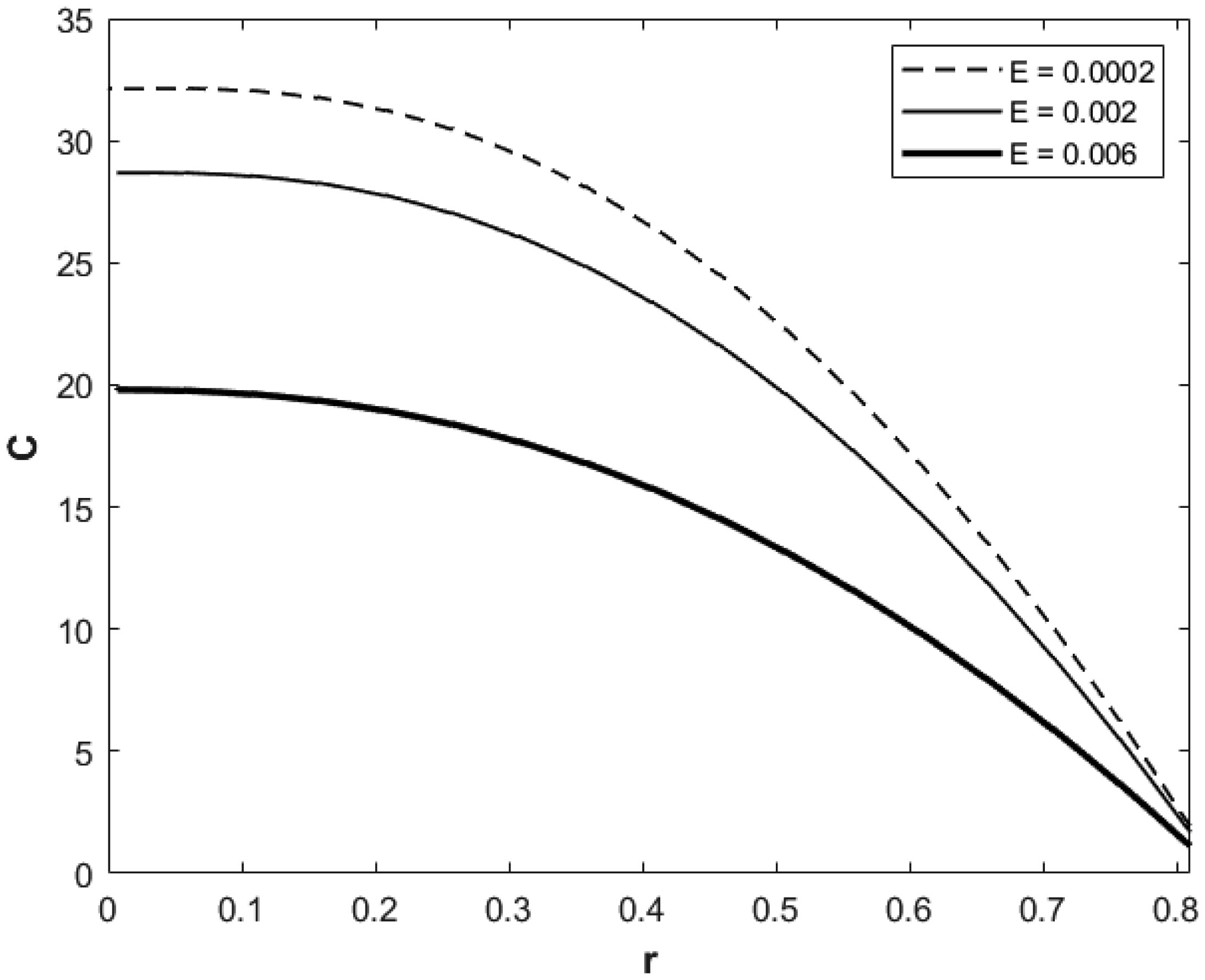
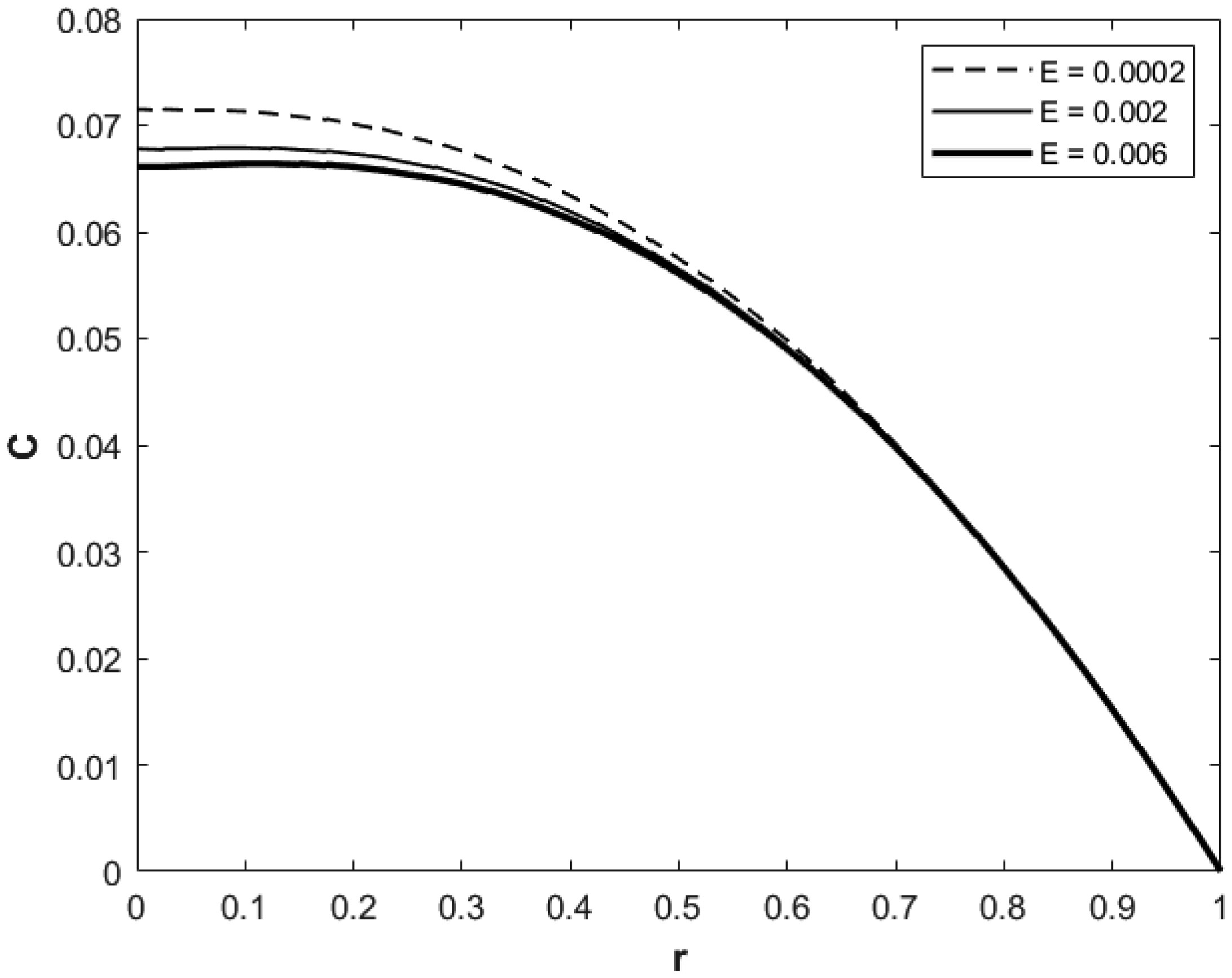
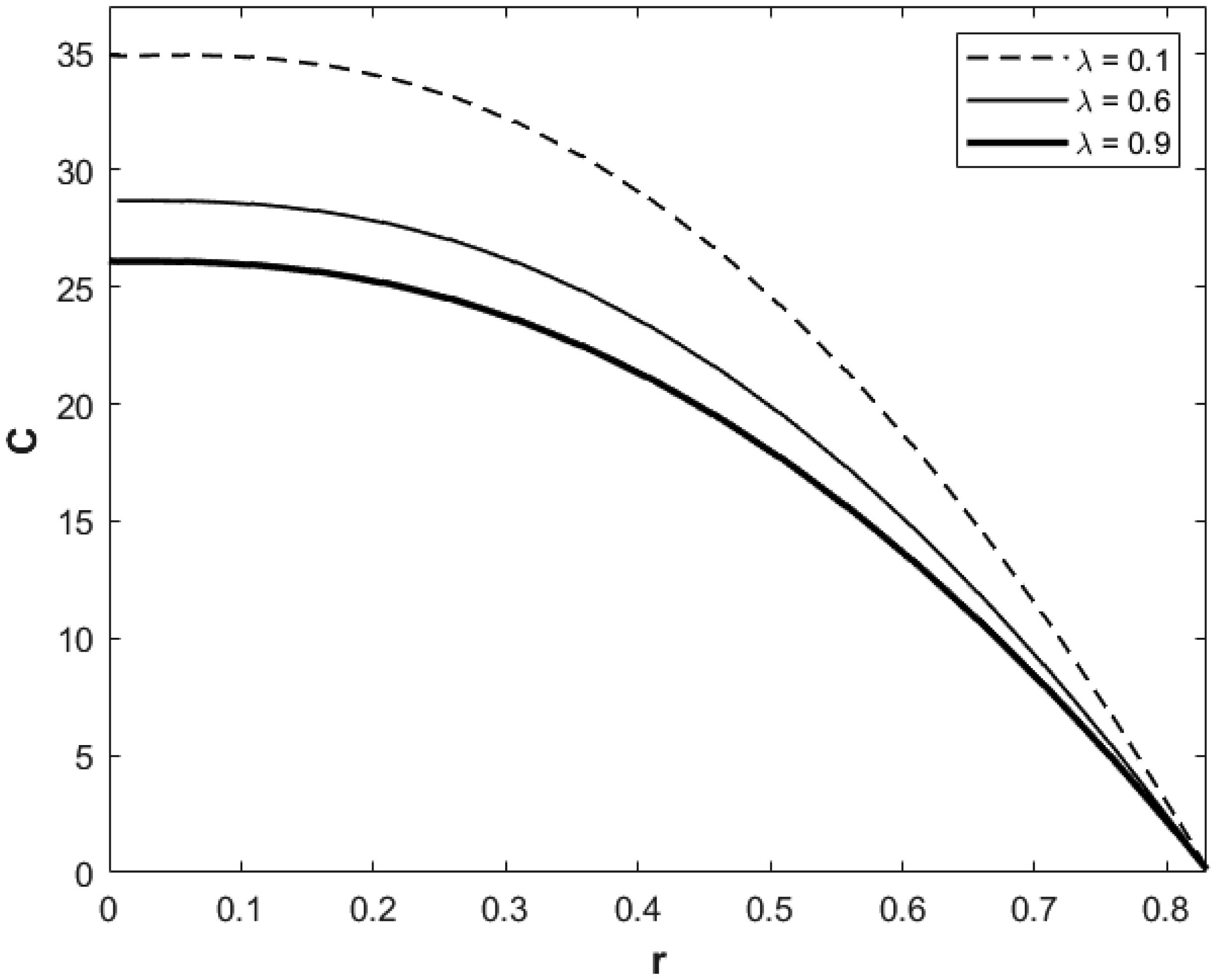
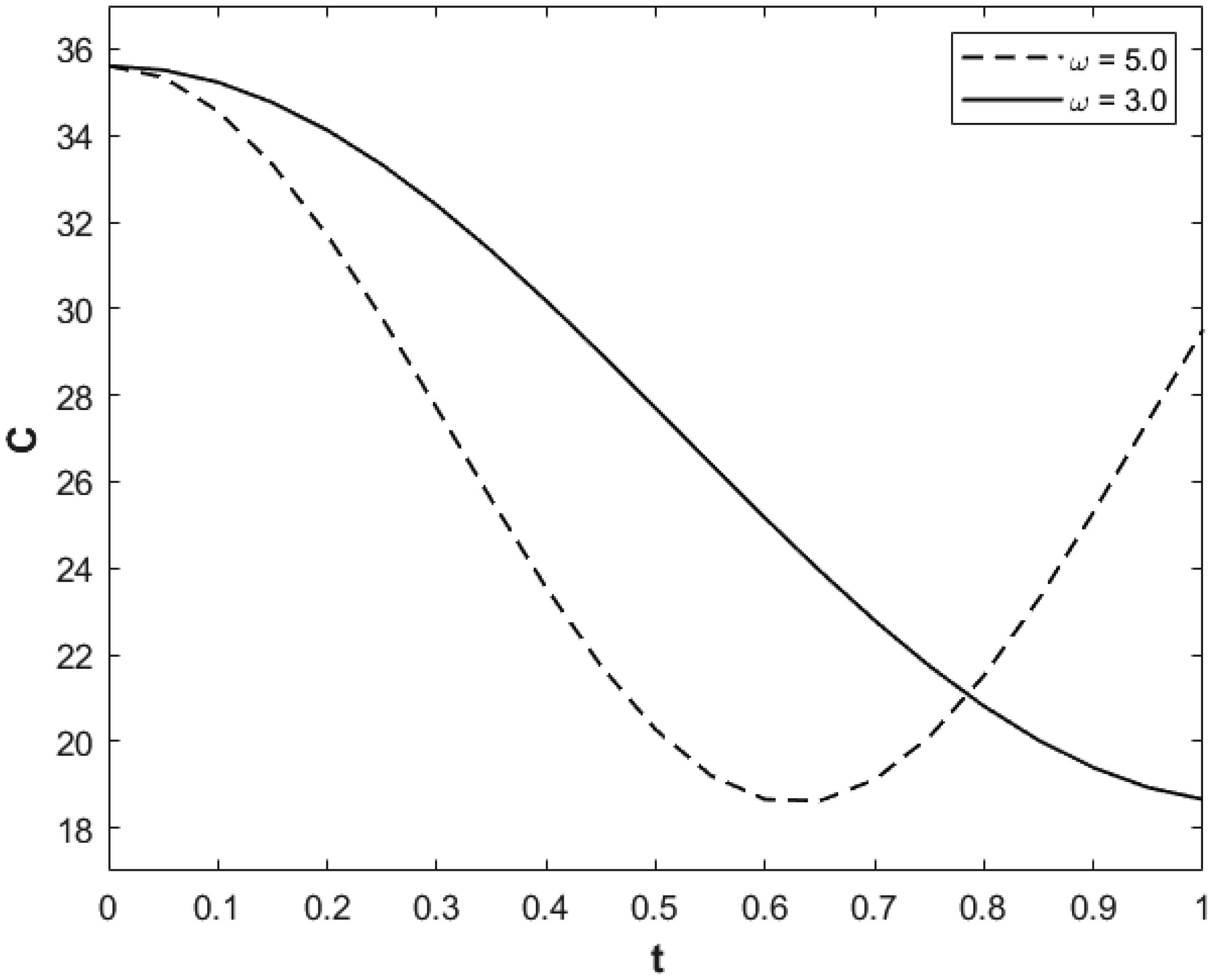
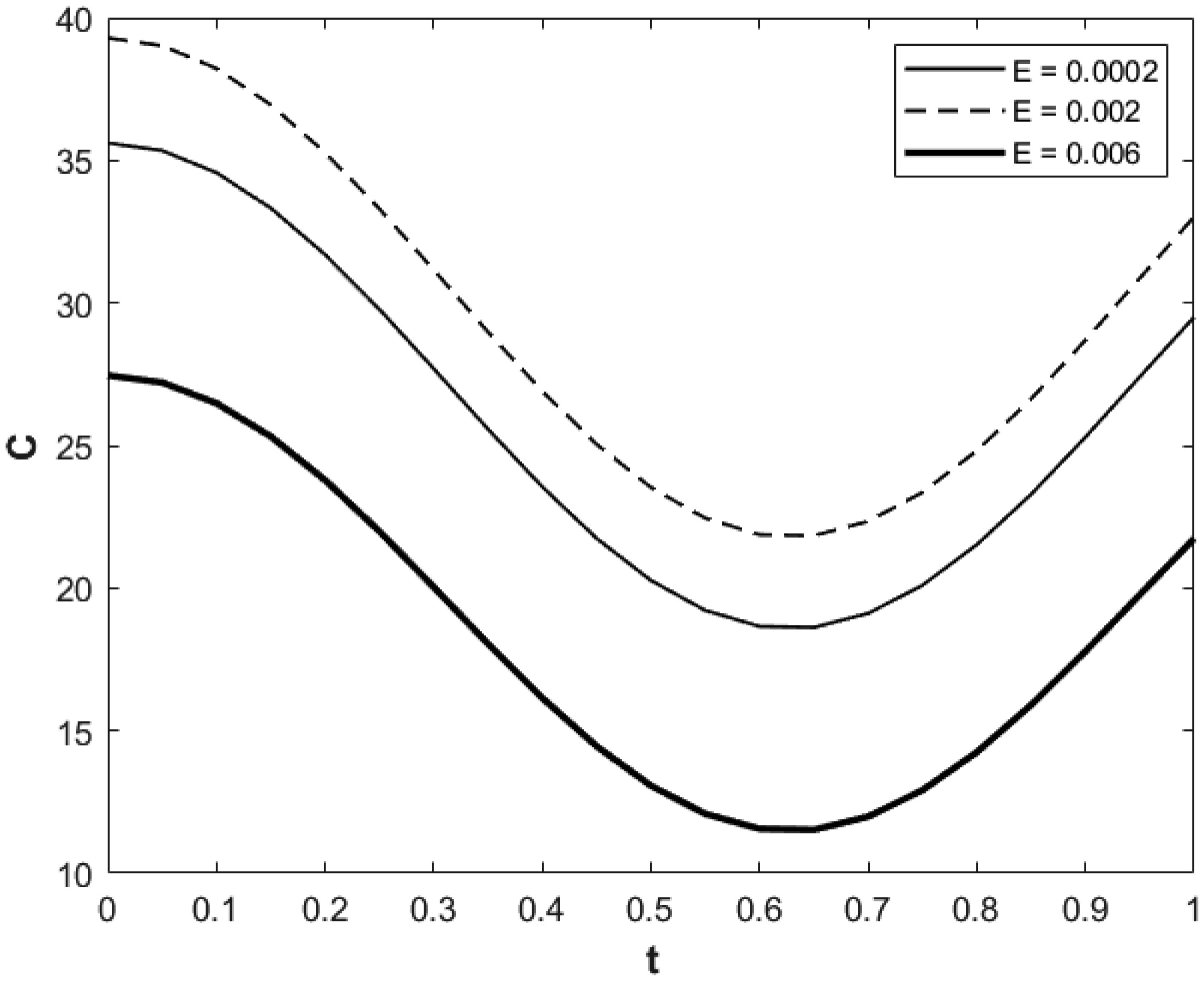
We investigated a physical system for unsteady blood flow and solute transport in a section of a constricted porous artery. The aim of this study was to determine effects of hematocrit, stenosis, pulse oscillation, diffusion, convection and chemical reaction on the solute transport. The significance of this study was uncovering combined roles played by stenosis height, hematocrit, pulse oscillation period, reactive rate, blood speed, blood pressure force and radial and axial extent of the porous artery on the solute transported by the blood flow in the described porous artery. We used both analytical and computational methods to determine blood flow quantities and solute transport for different parametric values of the described physical system. We found that solute transport increases with increasing stenosis height, blood pulsation period, convection and blood pressure force. However, transportation of solute reduces with increasing hematocrit, chemical reactive rate and radial or axial distance.
Citation: Daniel N. Riahi, Saulo Orizaga. Modeling and computation for unsteady blood flow and solute concentration in a constricted porous artery[J]. AIMS Bioengineering, 2023, 10(1): 67-88. doi: 10.3934/bioeng.2023007
We investigated a physical system for unsteady blood flow and solute transport in a section of a constricted porous artery. The aim of this study was to determine effects of hematocrit, stenosis, pulse oscillation, diffusion, convection and chemical reaction on the solute transport. The significance of this study was uncovering combined roles played by stenosis height, hematocrit, pulse oscillation period, reactive rate, blood speed, blood pressure force and radial and axial extent of the porous artery on the solute transported by the blood flow in the described porous artery. We used both analytical and computational methods to determine blood flow quantities and solute transport for different parametric values of the described physical system. We found that solute transport increases with increasing stenosis height, blood pulsation period, convection and blood pressure force. However, transportation of solute reduces with increasing hematocrit, chemical reactive rate and radial or axial distance.
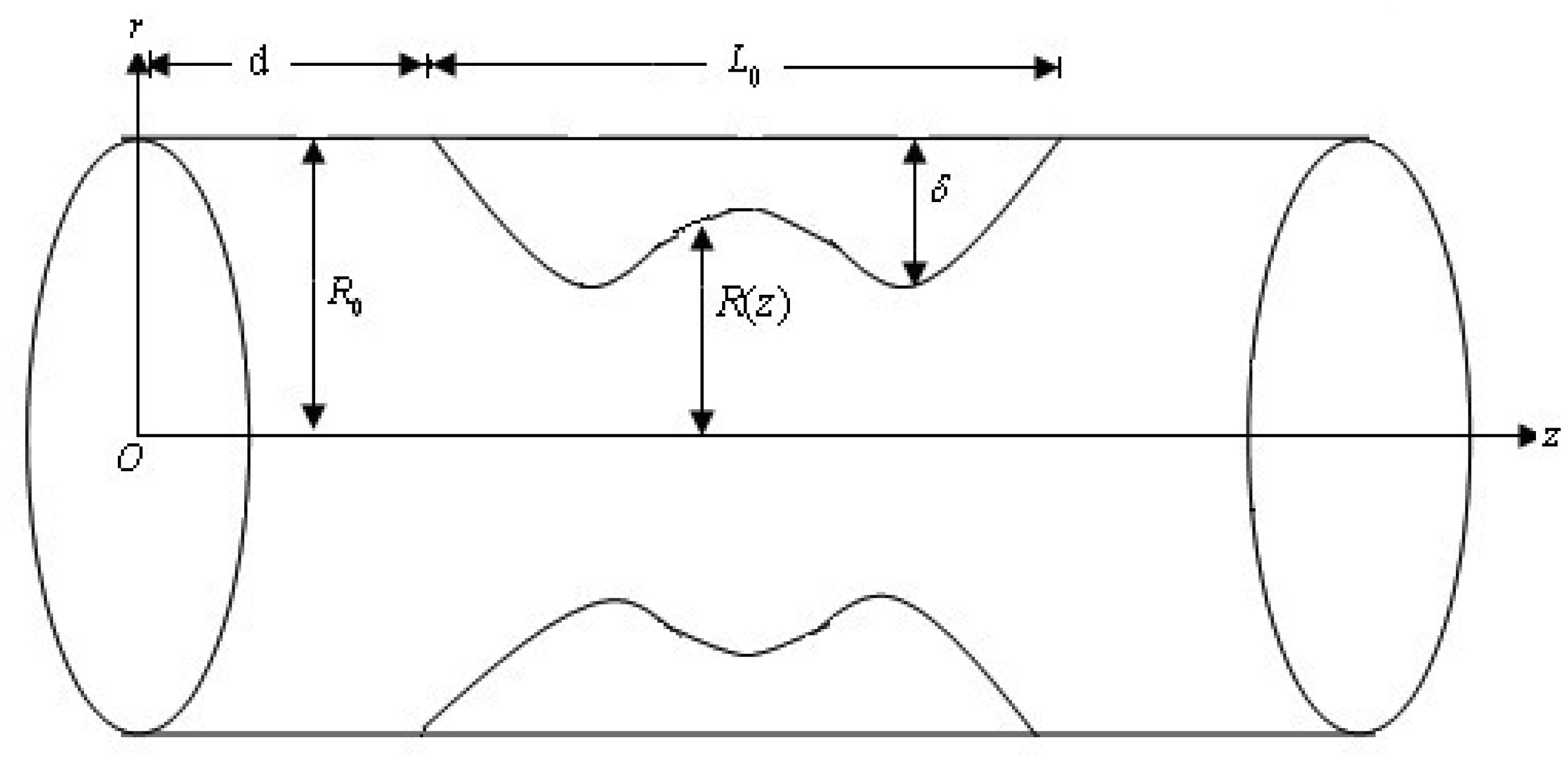
length of damaged stenotic region
location of stenosis
radial and axial coordinates, respectively
radius of stenotic region
radius of non-stenotic region
maximum radius of stenotic region
blood velocity vector along the axial direction
blood velocity vector along the radial direction
solute concentration
diffusivity coefficient
reaction constant
density
blood pressure
permeability of the porous medium
variable blood
dynamic viscosity of the plasma in the blood
maximum hematocrit at the center of cylindrical tube
parameter used for the formula's constriction
small parameter defined as the ratio of
flow resistance
wall shear stress of zeroth order
wall shear stress of 1st order

| [1] |
Srivastava LM, Srivastava VP (1983) On two-phase model of pulsatile blood flow with entrance effects. Biorheology 20: 761-777. https://doi.org/10.3233/BIR-1983-20604 
|
| [2] | Srivastava VP, Mishra S (2010) Non-Newtonian arterial blood flow through an overlapping stenosis. Appl Appl Math 5: 225-238. https://digitalcommons.pvamu.edu/aam/vol5/iss1/17 |
| [3] | Venkateswarlu K, Rao JA (2004) Numerical solution of unsteady blood flow through indented tube with atherosclerosis. Indian J Biochem Bio 41: 241-245. |
| [4] |
Back LH (1994) Estimated mean flow resistance during coronary artery catheterization. J Biomech 27: 169-175. https://doi.org/10.1016/0021-9290(94)90205-4
|
| [5] | Back LH, Kwack EY, Back MR (1996) Flow rate-pressure drop relation to coronary angioplasty: catheter obstruction effect. J Biomed Eng 118: 83-89. https://doi.org/10.1115/1.2795949 |
| [6] | Khanduri U, Sharma BK (2022) Hall and ion slip effects on hybrid nanoparticles (Au-GO/blood) flow through a catheterized stenosed artery with thrombosis. Proc Inst Mech Eng Part C 2022: 09544062221136710. https://doi.org/10.1177/09544062221136710 |
| [7] |
Saleem A, Akhtar S, Nadeem S, et al. (2021) Microphysical analysis for peristaltic flow of SWCNT and MWCNT carbon nanotubes inside a catheterised artery having thrombus: irreversibility effects with entropy. Int J Exergy 34: 301-314. https://doi.org/10.1504/IJEX.2021.113845
|
| [8] |
Srivastava VP, Rastogi R (2010) Blood flow through a stenosed catheterized artery: Effects of hematocrit and stenosis shape. Comput Math Appl 59: 1377-1385. https://doi.org/10.1016/j.camwa.2009.12.007
|
| [9] | Riahi DN (2016) Modeling unsteady two-phase blood flow in catheterized elastic artery with stenosis. Eng Sci Technol Int J 19: 1233-1243. https://doi.org/10.1016/j.jestch.2016.01.002 |
| [10] |
Srivastava VP (1996) Two-phase model of blood flow through stenosed tubes in the presence of a peripheral layer: applications. J Biomech 29: 1377-1382. https://doi.org/10.1016/0021-9290(96)00037-1
|
| [11] | Back LH, Cho YI, Crawford DW, et al. (1984) Effect of mild atherosclerosis on flow resistance in a coronary artery casting of man. Trans ASME 106: 48-53. https://doi.org/10.1115/1.3138456 |
| [12] |
Riahi DN (2017) On low frequency oscillatory elastic arterial blood flow with stenosis. Int J Appl Comput Math 3: 55-70. https://doi.org/10.1007/s40819-017-0341-5
|
| [13] |
Saleem S, Akhtar S, Nadeem S, et al. (2021) Mathematical study of electroosmotically driven peristaltic flow of Casson fluid inside a tube having systematically contracting and relaxing sinusoidal heated walls. Chinese J Phys 71: 300-311. https://doi.org/10.1016/j.cjph.2021.02.015
|
| [14] |
Khaled ARA, Vafai K (2003) The role of porous media in modeling flow and heat transfer in biological tissues. Int J Heat and Mass Transfer 46: 4989-5003. https://doi.org/10.1016/S0017-9310(03)00301-6
|
| [15] |
Lee DY, Vafai K (1999) Analytical characterization and conceptual assessment of solid and fluid temperature differentials in porous media. Int J Heat and Mass Transfer 42: 423-435. https://doi.org/10.1016/S0017-9310(99)00166-0
|
| [16] |
Mann KG, Nesheim ME, Church WR, et al. (1990) Surface-dependent reactions of the vitamin K-dependence enzyme complexes. Blood 76: 1-16. https://doi.org/10.1182/blood.V76.1.1.1
|
| [17] |
Orizaga S, Riahi DN, Soto JR (2020) Drug delivery in catheterized arterial blood flow with atherosclerosis. Results Appl Math 7: 100117. https://doi.org/10.1016/j.rinam.2020.100117
|
| [18] |
Ponalagusamy R, Murugan D, Priyadharshini S (2022) Effect of rheology of non-Newtonian fluid and chemical reaction on a dispersion of a solute and implication to blood flow. Int J Appl Comput Math 8: 109. https://doi.org/10.1007/s40819-022-01312-6
|
| [19] |
Rana J, Murthy P (2017) Unsteady solute dispersion in small blood vessel using a two-phase Casson model. Proc R Soc A 473: 20170427. https://doi.org/10.1098/rspa.2017.0427
|
| [20] |
Roy AK, Bég OA (2021) Mathematical modeling of unsteady solute dispersion in two fluids (micropolar-Newtonian) blood flow with bulk reaction. Int Commun Heat Mass Transfer 122: 105169. https://doi.org/10.1016/j.icheatmasstransfer.2021.105169
|
| [21] |
Valencia A, Villanueva M (2006) Unsteady flow and mass transfer in models of stenotic arteries considering fluid-structure interaction. Int Commun Heat Mass Transfer 33: 966-975. https://doi.org/10.1016/j.icheatmasstransfer.2006.05.006
|
| [22] |
Gandhi R, Sharma BK, Kumawat C, et al. (2022) Modeling and analysis of magnetic hybrid nanoparticle (Au-Al2O3/blood) based drug delivery through a bell-shaped occluded artery with joule heating, viscous dissipation and variable viscosity effects. Proc Inst Mech Eng Part E 236: 2024-2043. https://doi.org/10.1177/09544089221080273
|
| [23] |
Khanduri U, Sharma BK (2022) Entropy analysis for mhd flow subject to temperature-dependent viscosity and thermal conductivity. Nonlinear Dynamics and Applications: Proceedings of the ICNDA 2022 . Cham: Springer International Publishing 457-471. https://doi.org/10.1007/978-3-030-99792-2_38
|
| [24] |
Sharma BK, Kumawat C (2021) Impact of temperature dependent viscosity and thermal conductivity on MHD blood flow through a stretching surface with ohmic effect and chemical reaction. Nonlinear Eng 10: 255-271. https://doi.org/10.1515/nleng-2021-0020
|
| [25] | Sharma M, Sharma B, Tripathi B (2022) Radiation effect on MHD copper suspended nanofluid flow through a stenosed artery with temperature-dependent viscosity. Int J Nonlinear Anal Appl 13: 2573-2584. http://dx.doi.org/10.22075/ijnaa.2021.22438.2362 |
| [26] |
Zhao T, Khan MR, Chu Y, et al. (2021) Entropy generation approach with heat and mass transfer in magnetohydrodynamic stagnation point flow of a tangent hyperbolic nanofluid. Appl Math Mech 42: 1205-1218. https://doi.org/10.1007/s10483-021-2759-5
|
| [27] |
Chen Z, Ma X, Zhan H, et al. (2022) Experimental investigation of solute transport across transition interface of porous media under reversible flow direction. Ecotox Environ Safe 238: 111566. https://doi.org/10.1016/j.ecoenv.2022.113566
|
| [28] |
Berkowitz B, Cortis A, Dror I, et al. (2009) Laboratory experiments on dispersive transport across interfaces: The role of flow direction. Water Resour Res 45: W02201. https://doi.org/10.1029/2008WR007342
|
| [29] |
Murugan D, Roy AK, Ponalagusamy R, et al. (2022) Tracer dispersion due to pulsatile casson fluid flow in a circular tube with chemical reaction modulated by externally applied electromagnetic fields. Int J Appl Comput Math 8: 221. https://doi.org/10.1007/s40819-022-01412-3
|
| [30] |
Ndenda JP, Shaw S, Njagarah JBH (2023) Shear induced fractionalized dispersion during Magnetic Drug Targeting in a permeable microvessel. Colloids Surf B 221: 113001. https://doi.org/10.1016/j.colsurfb.2022.113001
|
| [31] | Roy AK, Bég OA (2021) Asymptotic study of unsteady mass transfer through a rigid artery with multiple irregular stenoses. Appl Math Comput 410: 126485. https://doi.org/10.1016/j.amc.2021.126485 |
| [32] |
Roy AK, Saha AK, Ponalagusamy R, et al. (2020) Mathematical model on magneto-hydrodynamic dispersion in a porous medium under the influence of bulk chemical reaction. Korea-Aust Rheol J 32: 287-299. https://doi.org/10.1007/s13367-020-0027-0
|
| [33] |
Sharma MK, Bansal K, Bansal S (2012) Pulsatile unsteady flow of blood through porous medium in a stenotic artery under the influence of transverse magnetic field. Korea-Aust Rheol J 24: 181-189. https://doi.org/10.1007/s13367-012-0022-1
|
| [34] |
Lapwood ER (1948) Convection of a fluid in a porous medium. Proceedings of the Cambridge Philosophical Society . UK: Cambridge University Press 508-521. https://doi.org/10.1017/S030500410002452X
|
| [35] | White FM (1991) Viscous Fluid Flow. New York: McGraw-Hill Inc. |
| [36] |
Sinha A, Shit GC (2015) Modeling of blood flow in a constricted porous vessel under magnetic environment: an analytical approach. Int J App Comput Math 1: 219-234. https://doi.org/10.1007/s40819-014-0022-6
|
| [37] |
Ascher U, Mathheij RM, Russell RD (1995) Numerical Solution of Boundary Value Problems for Ordinary Differential Equations. Philadelphia: SIAM Publication. https://doi.org/10.1137/1.9781611971231
|
Figures(11)

Daniel N. Riahi, Saulo Orizaga. Modeling and computation for unsteady blood flow and solute concentration in a constricted porous artery[J]. AIMS Bioengineering, 2023, 10(1): 67-88. doi: 10.3934/bioeng.2023007







 DownLoad:
DownLoad: