Citation: Poonam Sharma, Shalise Burch, Tejasvi Peesay, Susan M. Hamilla, Adam H. Hsieh, Carlos Luna Lopez. Downregulation of vimentin intermediate filaments affect human mesenchymal stem cell adhesion and formation of cellular projections[J]. AIMS Bioengineering, 2020, 7(4): 272-288. doi: 10.3934/bioeng.2020023

| [1] |
Jo CH, Lee YG, Shin WH, et al. (2014) Intra-articular injection of mesenchymal stem cells for the treatment of osteoarthritis of the knee: a proof-of-concept clinical trial. Stem Cells 32: 1254-1266. doi: 10.1002/stem.1634

|
| [2] |
Orozco L, Soler R, Morera C, et al. (2011) Intervertebral disc repair by autologous Mesenchymal bone marrow cells: a pilot study. Transplantation 92: 822-828. doi: 10.1097/TP.0b013e3182298a15
|
| [3] |
Squillaro T, Peluso G, Galderisi U (2016) Clinical trials with mesenchymal stem cells: an update. Cell Transplant 25: 829-848. doi: 10.3727/096368915X689622
|
| [4] | Kang SK, Shin IS, Ko MS, et al. (2012) Journey of mesenchymal stem cells for homing: strategies to enhance efficacy and safety of stem cell Therapy. Stem Cells Int 2012: e342968. |
| [5] |
Chapel A, Bertho JM, Bensidhoum M, et al. (2003) Mesenchymal stem cells home to injured tissues when co-infused with hematopoietic cells to treat a radiation-induced multi-organ failure syndrome. J Gene Med 5: 1028-1038. doi: 10.1002/jgm.452
|
| [6] |
Sasaki M, Abe R, Fujita Y, et al. (2008) Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J Immunol 180: 2581-2587. doi: 10.4049/jimmunol.180.4.2581
|
| [7] |
Wynn RF, Hart CA, Corradi-Perini C, et al. (2004) A small proportion of mesenchymal stem cells strongly expresses functionally active CXCR4 receptor capable of promoting migration to bone marrow. Blood 104: 2643-2645. doi: 10.1182/blood-2004-02-0526
|
| [8] |
Son B-R, Marquez-Curtis LA, Kucia M, et al. (2006) Migration of bone marrow and cord blood mesenchymal stem cells in vitro is regulated by stromal-derived factor-1-CXCR4 and hepatocyte growth factor-c-met axes and involves matrix metalloproteinases. Stem Cells 24: 1254-1264. doi: 10.1634/stemcells.2005-0271
|
| [9] |
McBeath R, Pirone DM, Nelson CM, et al. (2004) Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev Cell 6: 483-495. doi: 10.1016/S1534-5807(04)00075-9
|
| [10] |
Kilian KA, Bugarija B, Lahn BT, et al. (2010) Geometric cues for directing the differentiation of mesenchymal stem cells. Proc Natl Acad Sci 107: 4872-4877. doi: 10.1073/pnas.0903269107
|
| [11] |
Engler AJ, Sen S, Sweeney HL, et al. (2006) Matrix elasticity directs stem cell lineage specification. Cell 126: 677-689. doi: 10.1016/j.cell.2006.06.044
|
| [12] |
Curran JM, Chen R, Hunt JA (2006) The guidance of human mesenchymal stem cell differentiation in vitro by controlled modifications to the cell substrate. Biomaterials 27: 4783-4793. doi: 10.1016/j.biomaterials.2006.05.001
|
| [13] |
Salasznyk RM, Williams WA, Boskey A, et al. (2004) Adhesion to vitronectin and collagen I promotes osteogenic differentiation of human mesenchymal stem cells. J Biomed Biotechnol 2004: 24-34. doi: 10.1155/S1110724304306017
|
| [14] |
Ridley AJ, Schwartz MA, Burridge K, et al. (2003) Cell migration: integrating signals from front to back. Science 302: 1704-1709. doi: 10.1126/science.1092053
|
| [15] |
Schoumacher M, Goldman RD, Louvard D, et al. (2010) Actin, microtubules, and vimentin intermediate filaments cooperate for elongation of invadopodia. J Cell Biol 189: 541-556. doi: 10.1083/jcb.200909113
|
| [16] |
Havel LS, Kline ER, Salgueiro AM, et al. (2015) Vimentin regulates lung cancer cell adhesion through a VAV2–Rac1 pathway to control focal adhesion kinase activity. Oncogene 34: 1979-1990. doi: 10.1038/onc.2014.123
|
| [17] |
McInroy L, Määttä A (2007) Down-regulation of vimentin expression inhibits carcinoma cell migration and adhesion. Biochem Biophys Res Commun 360: 109-114. doi: 10.1016/j.bbrc.2007.06.036
|
| [18] |
Gonzales M, Weksler B, Tsuruta D, et al. (2001) Structure and function of a vimentin-associated matrix adhesion in endothelial cells. Mol Biol Cell 12: 85-100. doi: 10.1091/mbc.12.1.85
|
| [19] |
Tsuruta D (2003) The vimentin cytoskeleton regulates focal contact size and adhesion of endothelial cells subjected to shear stress. J Cell Sci 116: 4977-4984. doi: 10.1242/jcs.00823
|
| [20] |
Kreis S, Schönfeld H-J, Melchior C, et al. (2005) The intermediate filament protein vimentin binds specifically to a recombinant integrin α2/β1 cytoplasmic tail complex and co-localizes with native α2/β1 in endothelial cell focal adhesions. Exp Cell Res 305: 110-121. doi: 10.1016/j.yexcr.2004.12.023
|
| [21] |
Kim H, Nakamura F, Lee W, et al. (2010) Regulation of cell adhesion to collagen via β1 integrins is dependent on interactions of filamin A with vimentin and protein kinase C epsilon. Exp Cell Res 316: 1829-1844. doi: 10.1016/j.yexcr.2010.02.007
|
| [22] |
Kim H, Nakamura F, Lee W, et al. (2010) Filamin A is required for vimentin-mediated cell adhesion and spreading. Am J Physiol-Cell Physiol 298: C221-C236. doi: 10.1152/ajpcell.00323.2009
|
| [23] |
Bhattacharya R, Gonzalez AM, DeBiase PJ, et al. (2009) Recruitment of vimentin to the cell surface by β3 integrin and plectin mediates adhesion strength. J Cell Sci 122: 1390-1400. doi: 10.1242/jcs.043042
|
| [24] |
Homan SM, Martinez R, Benware A, et al. (2002) Regulation of the Association of α6β4 with vimentin intermediate filaments in endothelial cells. Exp Cell Res 281: 107-114. doi: 10.1006/excr.2002.5643
|
| [25] |
Liu C-Y, Lin H-H, Tang M-J, et al. (2015) Vimentin contributes to epithelial-mesenchymal transition cancer cell mechanics by mediating cytoskeletal organization and focal adhesion maturation. Oncotarget 6: 15966-15983. doi: 10.18632/oncotarget.3862
|
| [26] | Eckes B, Dogic D, Colucci-Guyon E, et al. (1998) Impaired mechanical stability, migration and contractile capacity in vimentin-deficient fibroblasts. J Cell Sci 111: 1897-1907. |
| [27] |
Bershadsky AD, Tint IS, Svitkina TM (1987) Association of intermediate filaments with vinculin-containing adhesion plaques of fibroblasts. Cell Motil Cytoskeleton 8: 274-283. doi: 10.1002/cm.970080308
|
| [28] |
Mendez MG, Kojima S-I, Goldman RD (2010) Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J 24: 1838-1851. doi: 10.1096/fj.09-151639
|
| [29] |
Burgstaller G, Gregor M, Winter L, et al. (2010) Keeping the vimentin network under control: cell–matrix adhesion–associated plectin 1f affects cell shape and polarity of fibroblasts. Mol Biol Cell 21: 3362-3375. doi: 10.1091/mbc.e10-02-0094
|
| [30] |
Huang L, Niu C, Willard B, et al. (2015) Proteomic analysis of porcine mesenchymal stem cells derived from bone marrow and umbilical cord: implication of the proteins involved in the higher migration capability of bone marrow mesenchymal stem cells. Stem Cell Res Ther 6: 7. doi: 10.1186/scrt541
|
| [31] |
Satelli A, Li S (2011) Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell Mol Life Sci 68: 3033-3046. doi: 10.1007/s00018-011-0735-1
|
| [32] |
Zhao Y, Yan Q, Long X, et al. (2008) Vimentin affects the mobility and invasiveness of prostate cancer cells. Cell Biochem Funct 26: 571-577. doi: 10.1002/cbf.1478
|
| [33] |
Helfand BT, Mendez MG, Murthy SNP, et al. (2011) Vimentin organization modulates the formation of lamellipodia. Mol Biol Cell 22: 1274-1289. doi: 10.1091/mbc.e10-08-0699
|
| [34] |
Menko AS, Bleaken BM, Libowitz AA, et al. (2014) A central role for vimentin in regulating repair function during healing of the lens epithelium. Mol Biol Cell 25: 776-790. doi: 10.1091/mbc.e12-12-0900
|
| [35] |
Nieminen M, Henttinen T, Merinen M, et al. (2006) Vimentin function in lymphocyte adhesion and transcellular migration. Nat Cell Biol 8: 156-162. doi: 10.1038/ncb1355
|
| [36] |
Barberis L, Pasquali C, Bertschy-Meier D, et al. (2009) Leukocyte transmigration is modulated by chemokine-mediated PI3Kγ-dependent phosphorylation of vimentin. Eur J Immunol 39: 1136-1146. doi: 10.1002/eji.200838884
|
| [37] |
Sharma P, Bolten ZT, Wagner DR, et al. (2017) Deformability of human mesenchymal stem cells is dependent on vimentin intermediate filaments. Ann Biomed Eng 45: 1365-1374. doi: 10.1007/s10439-016-1787-z
|
| [38] |
Jiu Y, Lehtimäki J, Tojkander S, et al. (2015) Bidirectional interplay between vimentin intermediate filaments and contractile actin stress fibers. Cell Rep 11: 1511-1518. doi: 10.1016/j.celrep.2015.05.008
|
| [39] |
Burgstaller G, Gregor M, Winter L, et al. (2010) Keeping the vimentin network under control: cell–matrix adhesion–associated plectin 1f affects cell shape and polarity of fibroblasts. Mol Biol Cell 21: 3362-3375. doi: 10.1091/mbc.e10-02-0094
|
| [40] |
Gregor M, Osmanagic-Myers S, Burgstaller G, et al. (2014) Mechanosensing through focal adhesion-anchored intermediate filaments. FASEB J 28: 715-729. doi: 10.1096/fj.13-231829
|
| [41] |
Bornheim R, Müller M, Reuter U, et al. (2008) A dominant vimentin mutant upregulates Hsp70 and the activity of the ubiquitin-proteasome system, and causes posterior cataracts in transgenic mice. J Cell Sci 121: 3737-3746. doi: 10.1242/jcs.030312
|
| [42] |
Uyama N, Zhao L, Rossen EV, et al. (2006) Hepatic stellate cells express synemin, a protein bridging intermediate filaments to focal adhesions. Gut 55: 1276-1289. doi: 10.1136/gut.2005.078865
|
| [43] |
Whipple RA, Balzer EM, Cho EH, et al. (2008) Vimentin filaments support extension of tubulin-based microtentacles in detached breast tumor cells. Cancer Res 68: 5678-5688. doi: 10.1158/0008-5472.CAN-07-6589
|
| [44] |
Boyne LJ, Fischer I, Shea TB (1996) Role of vimentin in early stages of neuritogenesis in cultured hippocampal neurons. Int J Dev Neurosci 14: 739-748. doi: 10.1016/S0736-5748(96)00053-6
|
| [45] |
Paul CD, Hung W-C, Wirtz D, et al. (2016) Engineered models of confined cell migration. Annu Rev Biomed Eng 18: 159-180. doi: 10.1146/annurev-bioeng-071114-040654
|
| [46] |
Smith H, Whittall C, Weksler B, et al. (2011) Chemokines stimulate bidirectional migration of human mesenchymal stem cells across bone marrow endothelial cells. Stem Cells Dev 21: 476-486. doi: 10.1089/scd.2011.0025
|
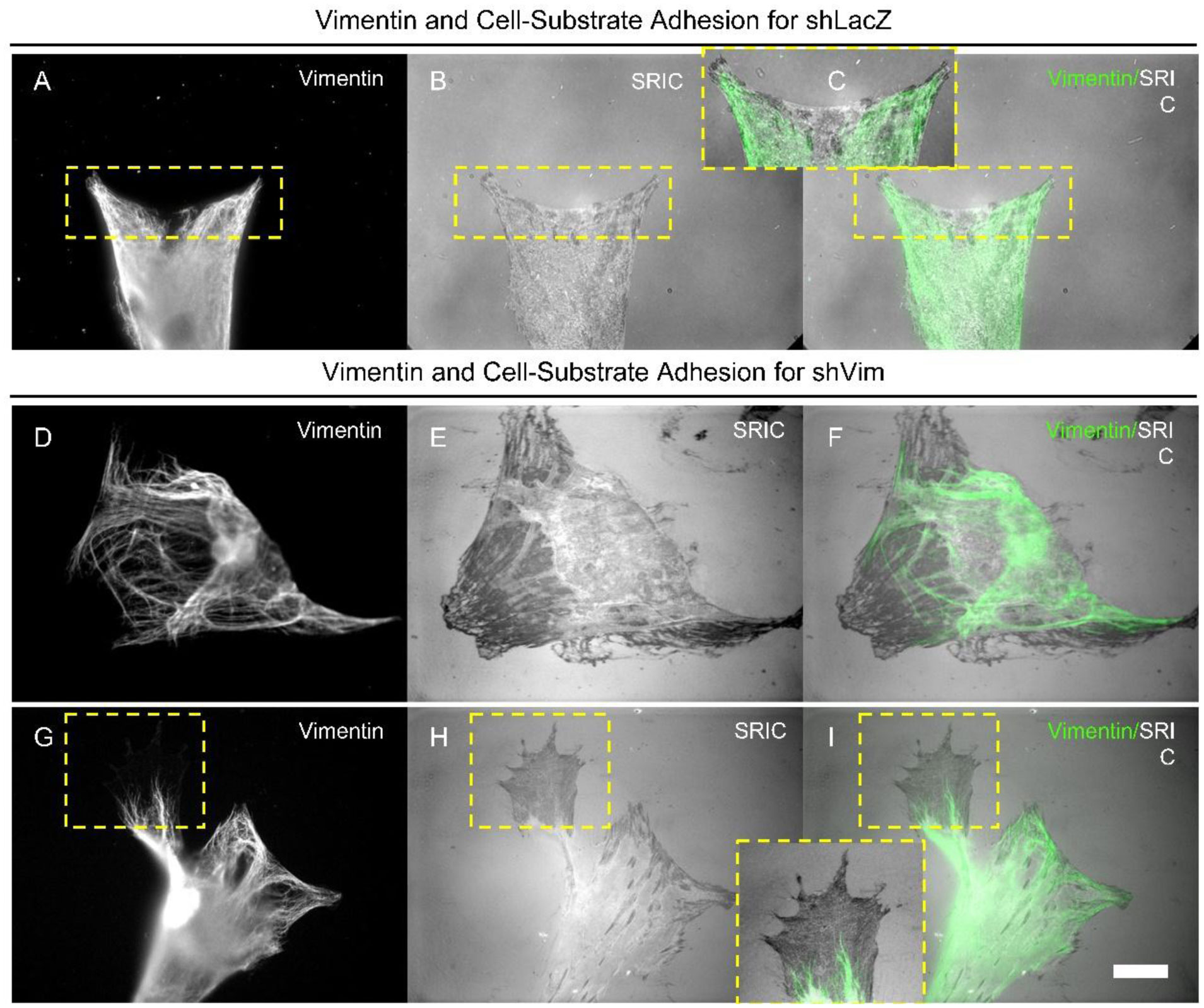
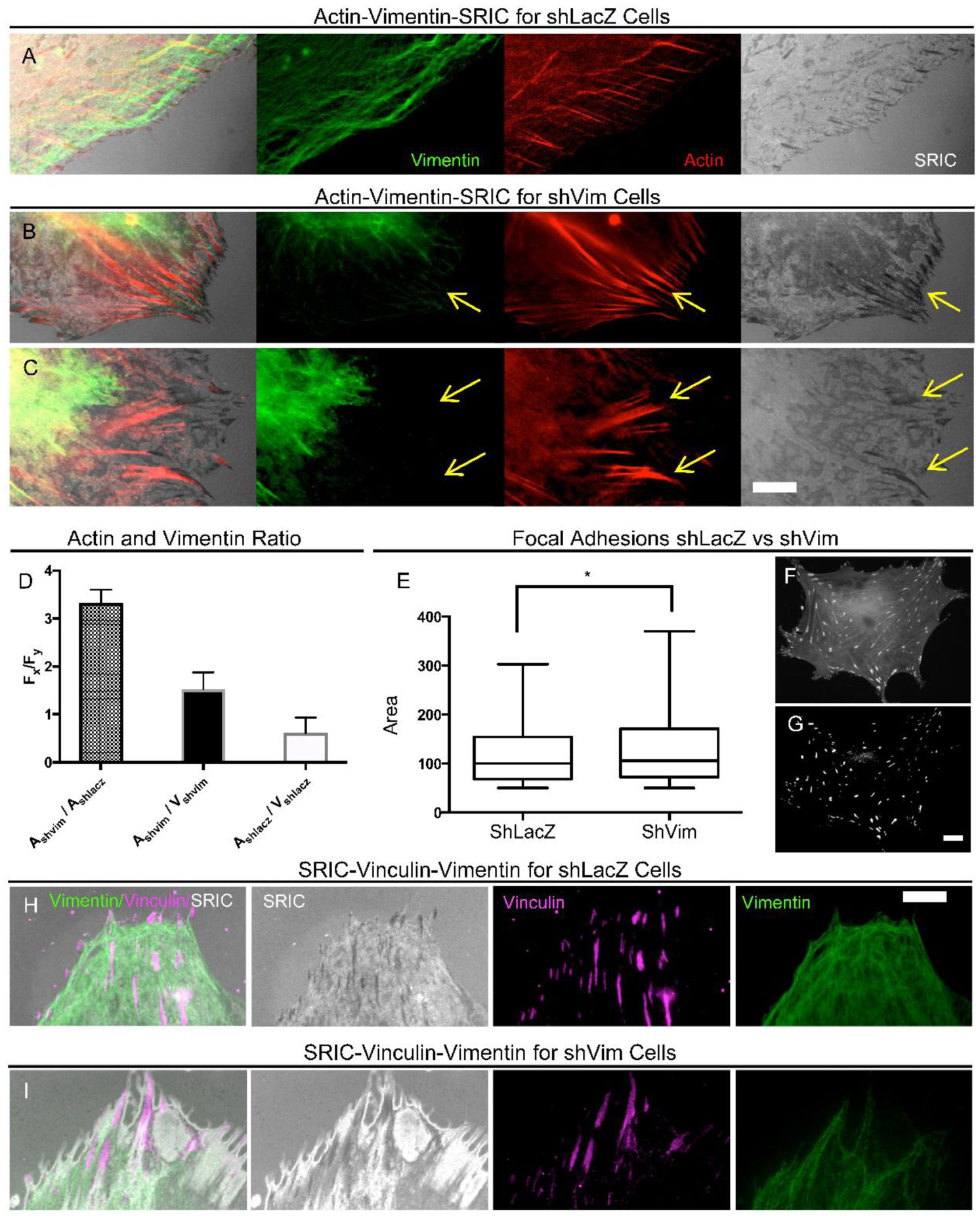
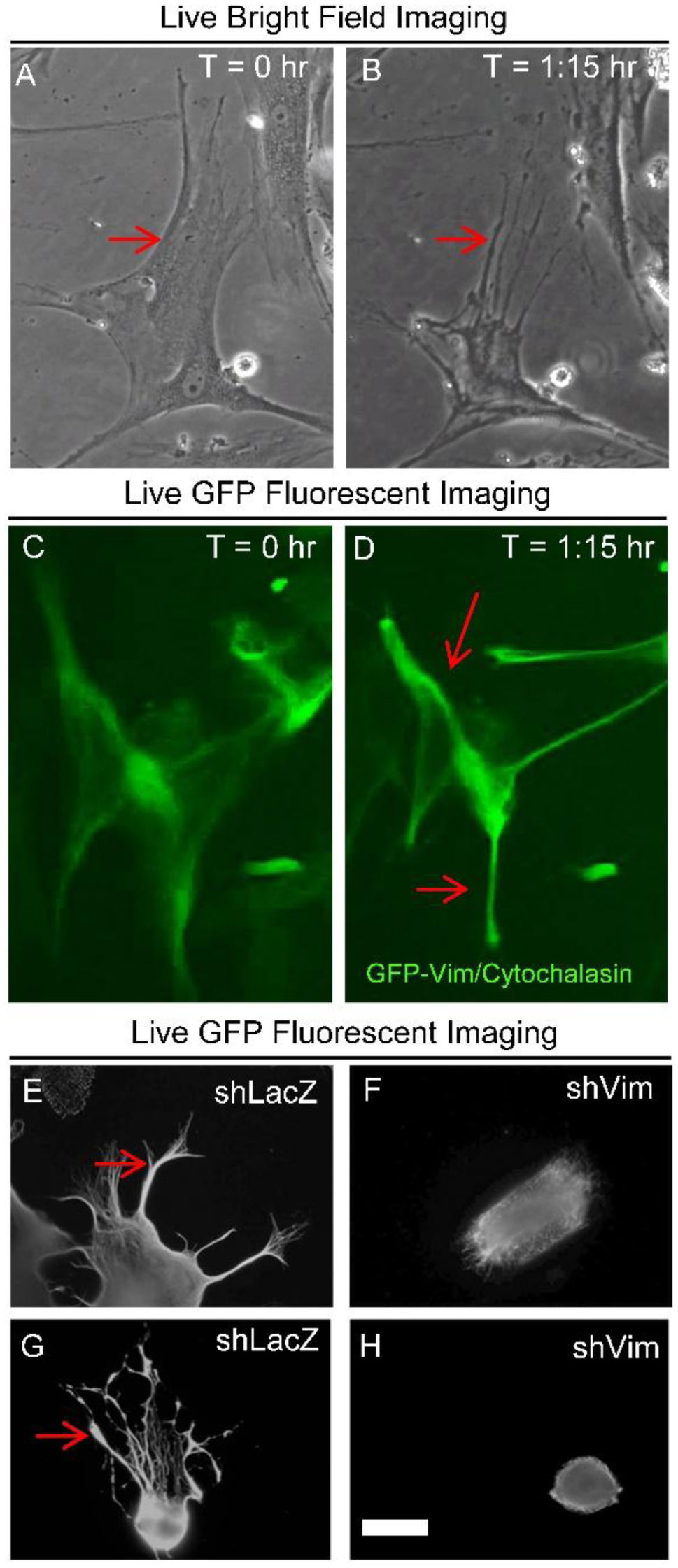
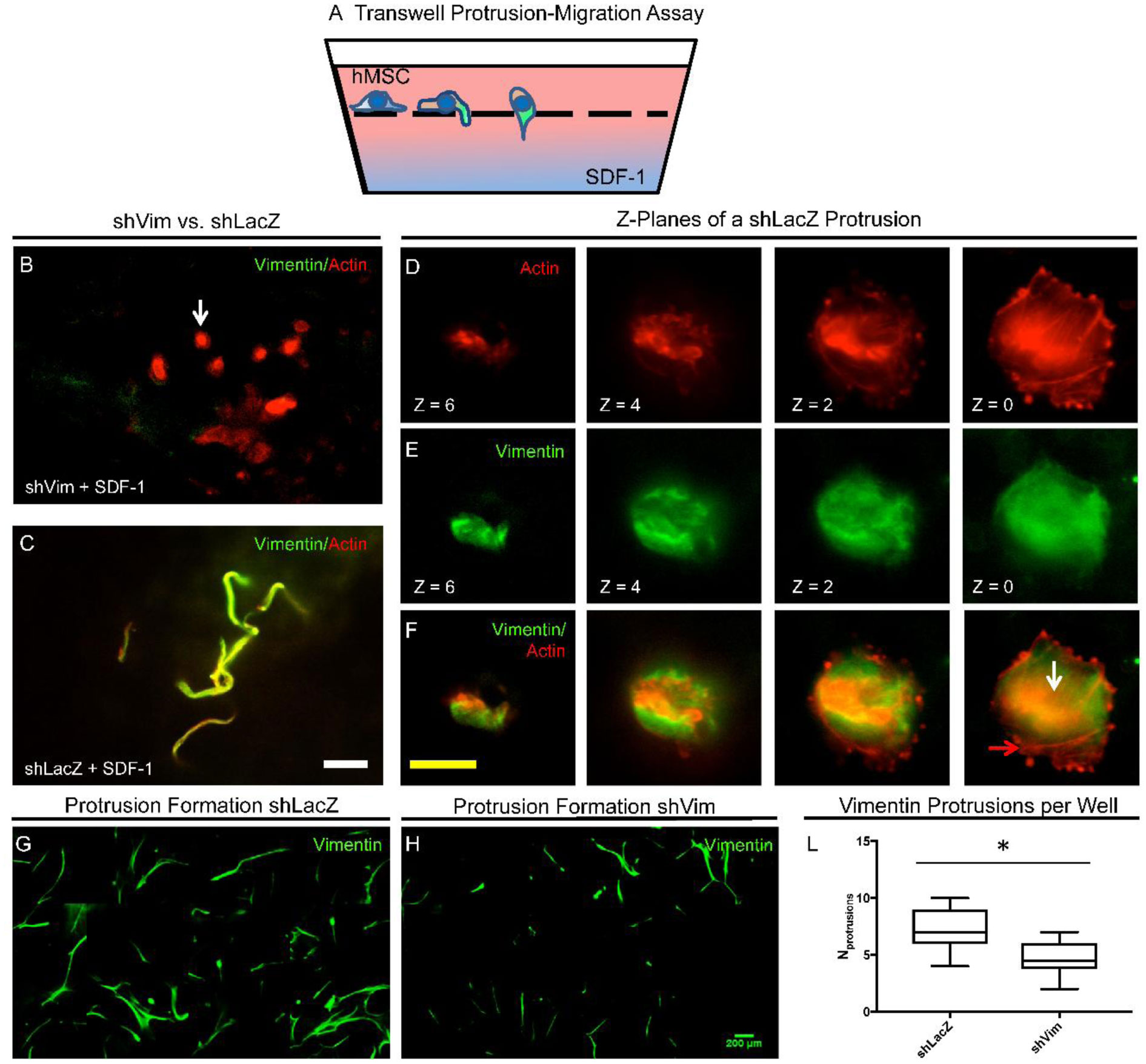
Figures(4)

Poonam Sharma, Shalise Burch, Tejasvi Peesay, Susan M. Hamilla, Adam H. Hsieh, Carlos Luna Lopez. Downregulation of vimentin intermediate filaments affect human mesenchymal stem cell adhesion and formation of cellular projections[J]. AIMS Bioengineering, 2020, 7(4): 272-288. doi: 10.3934/bioeng.2020023







 DownLoad:
DownLoad: