Citation: Junji Takaya. Small for Gestational Age and Magnesium: Intrauterine magnesium deficiency may induce metabolic syndrome in later life[J]. AIMS Public Health, 2015, 2(4): 793-803. doi: 10.3934/publichealth.2015.4.793

| [1] | [ Barker DJ, Hales CN, Fall CH. et al. (1993) Type 2 (non-insulin-dependent) diabetes mellitus, hypertension and hyperlipidaemia (syndrome X):relation to reduced fetal growth. Diabetologia 36: 62-67. |
| [2] | [ Hales CN, Barker DJ, Clark PM. et al. (1991) Fetal and infant growth and impaired glucose tolerance at age 64. BMJ 303:1019-1022. |
| [3] | [ Hales CN, Barker DJ. (2001) The thrifty phenotype hypothesis. Br Med Bull 60:5-20. |
| [4] | [ Lucas A. (1994) Programming in determing adult morbidity. Arch Dis Child 71: 288-290. |
| [5] | [ Gluckman PD, Hanson MA. (2004) Living with the past:'volution, development, and patterns of disease. Science 305:173-1736. |
| [6] | [ Lucas A. (1998) Programming by early nutrition: an experimental approach. J Nutr 128: suppl, 401S-406S. |
| [7] | [ Kanaka-Gantenbein C. (2010) Fetal origins of adult diabetes. Ann NY Acad Sci 1205:99-105. |
| [8] | [ Swanson JM, Entringer Buss SC, Wadhwa PD. (2009) Developmental origins of health and disease: environmental exposures. Semin Reprod Med 27: 391-402. |
| [9] | [ Stuefer S, Moncayo H, Moncayo R. (2015) The role of magnesium and thyroid function in early pregnancy after in-vitro fertilization (IVF): New aspects in endocrine physiology. BBA Clinical 3:196-204. |
| [10] | [ Barbosa NO, Okay TS, Loone CR. (2005) Magnesium and intrauterine growth restriction. J Am Coll Nutr 24:10-15. |
| [11] | [ Bullarbo M, Odman N, Nestler A. et al. (2013) Magnesium supplementation to prevent high blood pressure in pregnancy: a randomised placebo control trial. Arch Gynecol Obstet 288: 1269-1274. |
| [12] | [ Pathak P, Kapoor SK, Kapil U, Dwivdi SN. (2003) Serum magnesium level among pregnant women in a rural community of Haryana State, India. Eur J Clin Nutr 57:1504-1506. |
| [13] | [ Yang CY, Chiu HF, Tsai SS, Chang CC, Sung FC. (2002) Magnesium in drinking water and the risk of delivering a child of very low birth weight. Magnes Res 15:207-213. |
| [14] | [ Paolisso G, Scheen A, D'Onofrio, F, Lefebvre P. (1990) Magnesium and glucose homeostasis. Diabetologia 33: 511-514. |
| [15] | [ Rosolova H, Mayer O Jr, Reaven GM. (2000) Insulin-mediated glucose disposal is decreased in normal subjects with relatively low plasma magnesium concentrations. Metabolism 49: 418-420. |
| [16] | [ Resnick LM, Gupta RK, Gruenspan H, Alderman MH, Laragh JH. (1990) Hypertension and peripheral insulin resitance: possible mediating role of intracellular free magnesium. Am J Hypertens 3:373-379. |
| [17] | [ Nadler JL, Buchanan T, Natarajan R. et al. (1993) Magnesium deficiency produces insulin resistance and increased thromboxane synthesis. Hypertension 21:1024-1029. |
| [18] | [ Huerta MG, Roemmich JN, Kington ML. et al. (2005) Magnesium deficiency is associated with insulin resistance in obese children. Diabetes Care 28:1175-1181. |
| [19] | [ Lee PA, Chernausek SD, Hokken-Koelega AC, et al. (2003) International Small for Gestational Age Advisory Board consensus development conference statement: management of short children born for gestational age. Pediatrics 111:1253-1261. |
| [20] | [ Lall R, Wapnir RA. (2005) Meconium mineral content in small for gestational age neonates. Am J Perinatol 22: 259-263. |
| [21] | [ Husain ME, Mughal MZ. (1992) Mineral transport across the placenta. Arch Dis Child 67: 874-878. |
| [22] | [ Nandakumaran M, Dashti HM, Al-Zaid NS. (2002) Maternal-fetal transport kinetics of copper, selenium, magnesium and iron in perfused human placental lobule: in vitro study. Mol Cell Biochem 231: 9-14. |
| [23] | [ Standley PR, Standley CA. (2002) Identification of a functional Na+/Mg2+ exchanger in human trophoblast cells. Am J Hypertension 15: 565-570. |
| [24] | [ Husain SM, Mughal MZ, Sibley CP. (2000) Effects of acute maternal hyperglycaemia and hyperosmolarity on maternofetal transfer of calcium and magnesium across the in situ perfused rat placenta. Magnes Res 13: 239-247. |
| [25] | [ Yang H, Kim TH, Lee GS, Hong EJ, Jeung EB. (2014) Comparing the expression patterns of placental magnesium/phosphorus-transporting channels between healthy and preeclamptic pregnancies. Mol Reprod Dev 81:851-860. |
| [26] | [ Widdowson EM. (1981) Changes in body composition during growth. In: Davis JA, Dobbing J, eds. Scientific foundations of paediatrics. London: William Heinemann Medical Books Ltd, 330-342. |
| [27] | [ Handwerker SM, Altura BT, Jones KY, Altura BM. (1995) Maternal-fetal transfer of ionized serum magnesium during the stress of labor and delivery: a human study. J Am Coll Nutr 14: 376-381. |
| [28] | [ Bermudez L, Garcia-Vicent C, Lopez J, et al. (2015) Assessment of ten trace elements in umbilical cord blood and maternal blood: association with birth weight. J Transl Med 13:291. |
| [29] | [ Bischof P, Meisser A, Campana A. (2000) Paracrine and autocrine regulators of trophoblast invasion—a review. Placenta 21: Suppl A, Trophoblast Research 14: S55-S60. |
| [30] | [ Yi FX, Magness RR, Bird IM. (2005) Simultaneous imaging of [Ca2+]i and intracellular NO prodiuction in freshly isolated uterine artery endothelial cells: effect of ovarian cycle and pregnancy. Am J Physiol Regul Integr Comp Physiol 288: R140-R148. |
| [31] | [ Pearson PJ, Evora PRB, Seccombe JF. et al. (1998) Hypomagnesemia inhibits nitric oxide release from coronary endothelium: protective role of magnesium infusion after cardiac operations. Ann Thorac Surg 65: 967-972. |
| [32] | [ Lemancewicz A, Laudanska H, Laudanski T, Karpiuk A, Barta S. (2000) Permeability of fetal membranes to calcium and magnesium:possible role in preterm labour. Human Reproduction 15: 2018-2022. |
| [33] | [ Resnick LM, Gupta RK, Bhargava KK, et al. (1991) Cellular ions in hypertension, diabetes, and obesity. Hypertension 17: 951-957. |
| [34] | [ Trovati M, Anfossi G, Cavalot F, et al. (1988) Insulin directly reduces platelet sensitivity to aggregating agents. Diabetes 37: 780-786. |
| [35] | [ Takaya J, Higashino H, Kotera F, Kobayashi Y. (2003) Intracellular magnesium of platelets in children with diabetes and obesity. Metabolism 52: 468-471. |
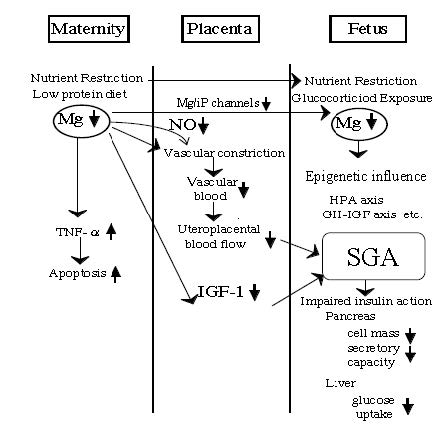
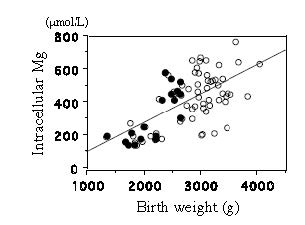
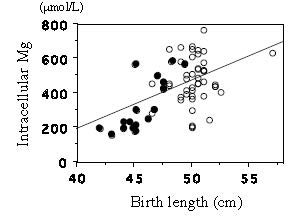
| [36] | [ Takaya J, Yamato F., Higashino H, Kobayashi Y. (2004) Relationship of intracellular magnesium of cord blood platelets to birth weight. Metabolism 53: 1544-1547. |
| [37] | [ Chiesa C, Osborn JF, Haass C, et al. (2008) Ghrelin, Leptin, IGF-1, IGFBP-3, and Insulin Concentrations at Birth: Is There a Relationship with Fetal Growth and Neonatal Anthropometry? Clin. Chem 54: 550-558. |
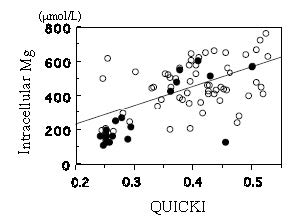
| [38] | [ Chen H, Sullivan G, Quon MJ. (2005) Assessing the Predictive Accuracy of QUICKI as a Surrogate Index for Insulin Sensitivity Using a Calibration Model. Diabetes 54: 1914-1925. |
| [39] | [ Takaya J, Yamato F, Higashino H, Kaneko K. (2007) Intracellular magnesium and adipokines in umbilical cord plasma and infant birth size. Pediatr Res 62: 700-703. |
| [40] | [ Phillips ID, Simonetta G, Owens JA, et al. (1996) Placental restriction alters has functional development of the pituitary-adrenal axis in the sheep fetus during late gestation. Pediatr Res 40: 861-866. |
| [41] | [ Ozanne SE, Smith GD, Tikerpae J, Hales CN. (1996) Altered regulation of hepatic glucose output in the male offspring of protein-malnourished rat dams. Am J Physiol 270: E559-E564. |
| [42] | [ Holt RI. (2002) Fetal programming of the growth hormone insulin-like growth factor axis. Trends. Endocrionol. Metabolism 13: 392-397. |
| [43] | [ Burdge GC, Hanson MA, Slater-Jefferies JL, Lillycrop KA. (2007) Epigenetic regulation of transcription: a mechanism for inducing variations in phenotype (fetal programming) by differences in nutrition during early life?" Br J Nutr 97:1036-1046. |
| [44] | [ Ross MG, Desai M. (2005) Gestational programming : population survival effects of drought and famine during pregnancy. Am J Physiol Regul Integr Comp Physiol 288: R25-R33. |
| [45] | [ Ozanne SE, Smith GD, Tikerpae J, Hales CN. (1996) Altered regulation of hepatic glucose output in the male offspring of protein-malnourished rat dams. Am J Physiol 270: E559-E564. |
| [46] | [ Hales CN, Barker DJ. (1992) Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty phenotype hypothesis. Diabetologia 35: 595-601. |
| [47] | [ Venu L, Kishore, DK, Raghnath M. (2005) Maternal and perinatal magnesium restriction predisposes rat pups to insulin resistance and glucose intolerance. J Nutr 135:1353-1358. |
| [48] | [ Reok W, Dean W, Walter J. (2001) Epigenetic reprogramming in mammalian development. Science 293:1089-1093. |
| [49] | [ Waterland RA, Jirtle RL. (2004) Early nutrition, epigenetic changes at transposons and imprinted genes, and enhanced susceptibility to adult chronic diseases. Nutrition 20:63-68. |
| [50] | [ De A, Ramesh V, Mahadevan S, Nagaraja V. (1998) Mg2+ mediated sequence-specific binding of transcriptional activator protein C. Biochemistry 37: 3831-3838. |
| [51] | [ Takaya J, Iharada A, Okihana H, Kaneko K. (2011) Magnesium deficiency in pregnant rats alters methylation of specific cytosines in the hepatic hydroxysteroid dehydrogenase-2 promoter of the offspring. Epigenetics 6:573-578. |
| [52] | [ Venu L, Raghunath M, Kishore YD, et al. (2008) Long-term effects of maternal magnesium restriction on adiposity and insulin resistance in rat pups. Obesity (Silver Spring) 16:1270-1276. |
| [53] | [ Altura BM, Shah NC, Jiang XC, et al. (2009) Short-term magnesium deficiency results in decreased levels of serum sphingomyelin, lipid peroxidation, and apoptosis in cardiovascular tissues. Am J Physiol Heart Circ Physiol 297:H86-92. |
| [54] | [ Shah NC, Shah GJ, Li Z, et al. (2014) Short-term magnesium deficiency downregulates telomerase, upregulates neutral sphingomyelinase and induces oxidative DNA damage in cardiovascular tissues: relevance to atherogenesis, cardiovascular diseases and aging. Int J Clin Exp Med 7:497-514. |
Figures(5) / Tables(1)

Junji Takaya. Small for Gestational Age and Magnesium: Intrauterine magnesium deficiency may induce metabolic syndrome in later life[J]. AIMS Public Health, 2015, 2(4): 793-803. doi: 10.3934/publichealth.2015.4.793







 DownLoad:
DownLoad: