Citation: Jianquan Li, Xiaoqin Wang, Xiaolin Lin. Impact of behavioral change on the epidemic characteristics of an epidemic model without vital dynamics[J]. Mathematical Biosciences and Engineering, 2018, 15(6): 1425-1434. doi: 10.3934/mbe.2018065

| [1] | [ F. Brauer, The Kermack-McKendrick epidemic model revisited, Math. Bios., 198 (2005), 119-131. |
| [2] | [ F. Brauer and C. Castillo-Chávez, Mathematical Models in Population Biology and Epidemiology, 2nd edn. Springer, New York, 2001. |
| [3] | [ V. Capasso and G. Serio, A generalizaition of the Kermack-McKendrick deterministic epidemic model, Math. Bios., 42 (1978), 43-61. |
| [4] | [ Y. H. Hsieh and C. W. S. Chen, Turning points, reproduction number, and impact of climatological events for multi-wave dengue outbreaks, Trop. Med. Int. Heal., 14 (2009), 628-638. |
| [5] | [ W. O. Kermack and A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. R. Soc. Lond. A, 115 (1927), 700-721. |
| [6] | [ J. Li, Y. Li and Y. Yang, Epidemic characteristics of two classic models and the dependence on the initial conditions, Math. Bios. Eng., 13 (2016), 999-1010. |
| [7] | [ J. Li and Y. Lou, Characteristics of an epidemic outbreak with a large initial infection size, J. Biol. Dyn., 10 (2016), 366-378. |
| [8] | [ J. Ma and D. J. D. Earn, Generality of the final size formula for an epidemic of a newly invading infectious disease, Bull. Math. Biol., 68 (2006), 679-702. |
| [9] | [ Z. Ma and J. Li, Dynamical Modeling and Analysis of Epidemics, Singapore, 2009. |
| [10] | [ J. C. Miller, A note on the derivation of epidemic final sizes, Bull. Math. Biol., 74 (2012), 2125-2141. |
| [11] | [ F. Zhang, J. Li and J. Li, Epidemic characteristics of two classic SIS models with disease-induced death, J. Theoret. Biol., 424 (2017), 73-83. |
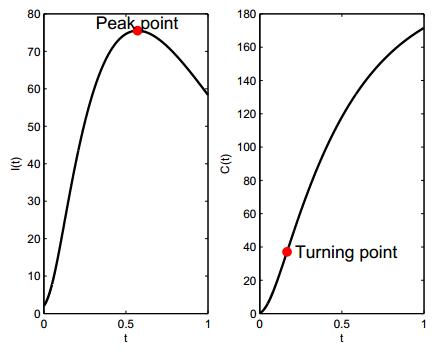
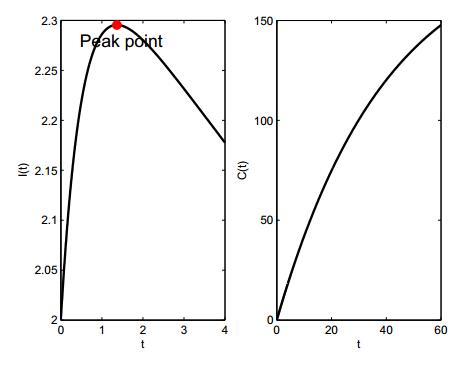
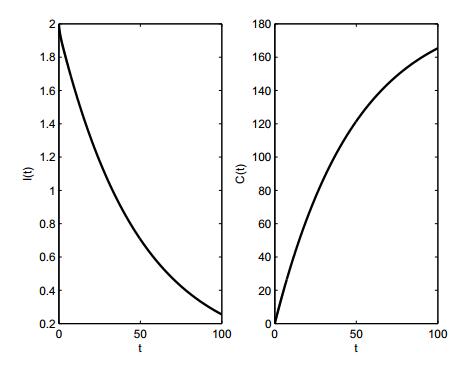
Figures(3)

Jianquan Li, Xiaoqin Wang, Xiaolin Lin. Impact of behavioral change on the epidemic characteristics of an epidemic model without vital dynamics[J]. Mathematical Biosciences and Engineering, 2018, 15(6): 1425-1434. doi: 10.3934/mbe.2018065







 DownLoad:
DownLoad: