Citation: Anna V. Babii, Anna L. Arkhipova, Irina N. Andreichenko, Artyom V. Brigida, Svetlana N. Kovalchuk. A TaqMan PCR assay for detection of DGAT1 K232A polymorphism in cattle[J]. AIMS Agriculture and Food, 2018, 3(3): 306-312. doi: 10.3934/agrfood.2018.3.306

| [1] |
Farese RV, Cases S, Smith SJ (2000) Triglyceride synthesis: Insights from the cloning of diacylglycerol acyltransferase. Curr Opin Lipidol 11: 229–234. doi: 10.1097/00041433-200006000-00002

|
| [2] |
Homa ST, Racowsky C, Mcgaughey RW (1986) Lipid analysis of immature pig oocytes. J Reprod Fertil 77: 425–434. doi: 10.1530/jrf.0.0770425
|
| [3] |
Cases S, Smith SJ, Zheng YW, et al. (1998) Identification of a gene encoding an acyl CoA: Diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc Natl Acad Sci 95: 13018–13023. doi: 10.1073/pnas.95.22.13018
|
| [4] |
Chen HC, Smith SJ, Ladha Z, et al. (2002) Increased insulin and leptin sensitivity in mice lacking acyl CoA: Diacylglycerol acyltransferase 1. J Clin Invest 109: 1049–1055. doi: 10.1172/JCI0214672
|
| [5] |
Smith SJ, Cases S, Jensen DR, et al. (2000) Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nat Genet 25: 87–90. doi: 10.1038/75651
|
| [6] |
Riquet J, Coppieters W, Cambisano N, et al. (1999) Fine-mapping of quantitative trait loci by identity by descent in outbred populations: Application to milk production in dairy cattle. Proc Natl Acad Sci 96: 9252–9257. doi: 10.1073/pnas.96.16.9252
|
| [7] | Farnir F, Grisart B, Coppieters W, et al. (2002) Simultaneous mining of linkage and linkage disequilibrium to fine map quantitative trait loci in outbred half-sib pedigrees: Revisiting the location of a quantitative trait locus with major effect on milk production on bovine chromosome 14. Genetics 161: 275–287. |
| [8] |
Winter A, Kramer W, Werner FAO, et al. (2002) Association of a lysine-232/alanine polymorphism in a bovine gene encoding acyl-CoA: Diacylglycerol acyltransferase (DGAT1) with variation at a quantitative trait locus for milk fat content. Proc Natl Acad Sci 99: 9300–9305. doi: 10.1073/pnas.142293799
|
| [9] |
Grisart B, Coppieters W, Farnir F, et al. (2002) Positional candidate cloning of a QTL in dairy cattle: Identification of a missense mutation in the bovine DGAT1 gene with major effect on milk yield and composition. Genome Res 12: 222–231. doi: 10.1101/gr.224202
|
| [10] |
Kong HS, Oh JD, Lee JH, et al. (2007) Association of sequence variations in DGAT 1 gene with economic traits in hanwoo (Korea Cattle). Asian-Australas J Anim Sci 20: 817–820. doi: 10.5713/ajas.2007.817
|
| [11] |
Rosse IDC, Steinberg RDS, Coimbra RS, et al. (2014) Novel SNPs and INDEL polymorphisms in the 3′UTR of DGAT1 gene: In silico analyses and a possible association. Mol Biol Rep 41: 4555–4563. doi: 10.1007/s11033-014-3326-z
|
| [12] |
Spelman RJ, Ford CA, Mcelhinney P, et al. (2002) Characterization of the DGAT1 Gene in the New Zealand Dairy Population. J Dairy Sci 85: 3514–3517. doi: 10.3168/jds.S0022-0302(02)74440-8
|
| [13] |
Fisher PJ, Spelman RJ (2004) Verification of selective DNA pooling methodology through identification and estimation of the DGAT1 effect. Anim Genet 35: 201–205. doi: 10.1111/j.1365-2052.2004.01137.x
|
| [14] |
Tăbăran A, Balteanu VA, Gal E, et al. (2015) Influence of DGAT1 K232A polymorphism on milk fat percentage and fatty acid profiles in Romanian Holstein Cattle. Anim Biotechnol 26: 105–111. doi: 10.1080/10495398.2014.933740
|
| [15] |
Thaller G, Kühn C, Winter A, et al. (2003) DGAT1, a new positional and functional candidate gene for intramuscular fat deposition in cattle. Anim Genet 34: 354–357. doi: 10.1046/j.1365-2052.2003.01011.x
|
| [16] |
Avilés C, Polvillo O, Peña F, et al. (2013) Associations between DGAT1, FABP4, LEP, RORC, and SCD1 gene polymorphisms and fat deposition in Spanish commercial beef. J Anim Sci 91: 4571–4577. doi: 10.2527/jas.2013-6402
|
| [17] | Gene Runner, 2017. Available from: http://www.generunner.net/. |
| [18] | Multiple Primer Analyzer, 2017. Available from: |
| [19] | https://www.thermofisher.com/in/en/home/brands/thermo-scientific/molecular-biology/molecular-biology-learning-center/molecular-biology-resource-library/thermo-scientific-web-tools/multiple-primer-analyzer.html. |
| [20] |
19. Glazko VI, Andreichenko IN, Kovalchuk SN, et al. (6) Candidate genes for control of cattle milk production traits. Russ Agric Sci 42: 458–464. doi: 10.3103/S1068367416060082
|
| [21] | 20. Komisarek J, Michalak A (2008) A relationship between DGAT1 K232A polymorphism and selected reproductive traits in Polish Holstein-Friesian cattle. Anim Sci Pap Rep 26: 89–95. |
| [22] |
21. Abdolmohammadi A, Atashi H, Zamani P, et al. (2011) High resolution melting as an alternative method to genotype diacylglycerol O-acyltransferase 1 (DGAT1) K232A polymorphism in cattle. Czech J Anim Sci 56: 370–376. doi: 10.17221/2393-CJAS
|
| [23] |
22. Rashydov AN, Spiridonov VG, Konoval ON, et al. (2010) Identification of allele variants of cattle milk productivity genes using PCR and the anti-primer method. Cytol Genet 44: 272–275. doi: 10.3103/S0095452710050026
|
| [24] |
23. Näslund J, Fikse WF, Pielberg GR, et al. (2008) Frequency and Effect of the Bovine Acyl-CoA: Diacylglycerol Acyltransferase 1 (DGAT1) K232A Polymorphism in Swedish Dairy Cattle. J Dairy Sci 91: 2127–2134. doi: 10.3168/jds.2007-0330
|
| [25] | 24. Berg H (2012) Restriction Fragment Length Polymorphism Analysis of PCR-Amplified Fragments (PCR-RFLP) and Gel Electrophoresis-Valuable Tool for Genotyping and Genetic Fingerprinting, In: Gel Electrophoresis-Principles and Basics, IntechOpen, 315–334. |
| [26] |
25. Herrmann MG, Durtschi JD, Bromley LK, et al. (2006) Amplicon DNA melting analysis for mutation scanning and genotyping: Cross-platform comparison of instruments and dyes. Clin Chem 52: 494–503. doi: 10.1373/clinchem.2005.063438
|
| [27] |
26. Seipp MT, Durtschi JD, Liew MA, et al. (2007) Unlabeled oligonucleotides as internal temperature controls for genotyping by amplicon melting. J Mol Diagn 9: 284–289. doi: 10.2353/jmoldx.2007.060136
|
| [28] |
27. Krypuy M, Newnham GM, Thomas DM, et al. (2006) High resolution melting analysis for the rapid and sensitive detection of mutations in clinical samples: KRAS codon 12 and 13 mutations in non-small cell lung cancer. BMC Cancer 6: 295. doi: 10.1186/1471-2407-6-295
|
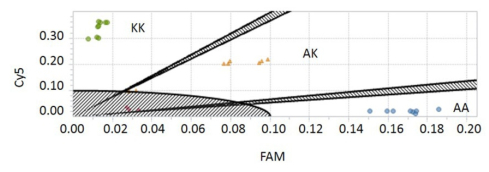
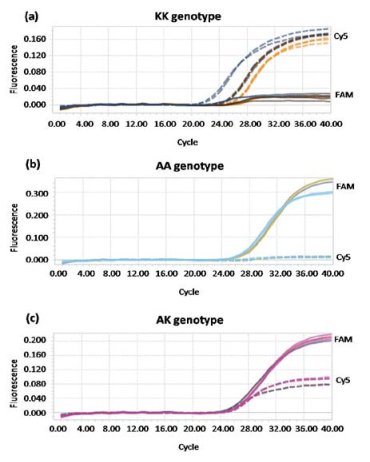
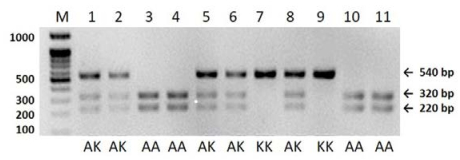
Figures(3) / Tables(1)

Anna V. Babii, Anna L. Arkhipova, Irina N. Andreichenko, Artyom V. Brigida, Svetlana N. Kovalchuk. A TaqMan PCR assay for detection of DGAT1 K232A polymorphism in cattle[J]. AIMS Agriculture and Food, 2018, 3(3): 306-312. doi: 10.3934/agrfood.2018.3.306







 DownLoad:
DownLoad: