Citation: Ezekiel Mugendi Njeru. Exploiting diversity to promote arbuscular mycorrhizal symbiosis and crop productivity in organic farming systems[J]. AIMS Agriculture and Food, 2018, 3(3): 280-294. doi: 10.3934/agrfood.2018.3.280

| [1] |
Godfray HCJ, Beddington JR, Crute IR, et al. (2010) Food security: The challenge of feeding 9 billion people. Science 327: 812–818. doi: 10.1126/science.1185383

|
| [2] |
Bainard L, Klironomos J, Gordon A (2011) The mycorrhizal status and colonization of 26 tree species growing in urban and rural environments. Mycorrhiza 21: 91–96. doi: 10.1007/s00572-010-0314-6
|
| [3] |
Tilman D (1998) The greening of the green revolution. Nature 396: 211–212. doi: 10.1038/24254
|
| [4] |
Schüßler A, Schwarzott D, Walker C (2001) A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol Res 105: 1413–1421. doi: 10.1017/S0953756201005196
|
| [5] |
Fitter AH (2005) Darkness visible: Reflections on underground ecology. J Ecol 93: 231–243. doi: 10.1111/j.0022-0477.2005.00990.x
|
| [6] |
Wang B, Qiu YL (2006) Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 16: 299–363. doi: 10.1007/s00572-005-0033-6
|
| [7] |
Redecker D, Kodner R, Graham LE (2000) Glomalean fungi from the Ordovician. Science 289: 1920–1921. doi: 10.1126/science.289.5486.1920
|
| [8] |
Krings M, Taylor TN, Hass H, et al. (2007) Fungal endophytes in a 400-million-yr-old land plant: Infection pathways, spatial distribution, and host responses. New Phytol 174: 648–657. doi: 10.1111/j.1469-8137.2007.02008.x
|
| [9] |
Parniske M (2008) Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat Rev Micro 6: 763–775. doi: 10.1038/nrmicro1987
|
| [10] |
Pandey M, Sharma J, Taylor DL, et al. (2013) A narrowly endemic photosynthetic orchid is non-specific in its mycorrhizal associations. Mol Ecol 22: 2341–2354. doi: 10.1111/mec.12249
|
| [11] | Willer H, Lernoud J (2017) The world of organic agriculture-statistics and emerging trends 2017, Research Institute of Organic Agriculture FiBL. |
| [12] | Willer H, Kilcher L (2012) (Eds.) The world of organic agriculture-statistics and emerging trends 2012, Research Institute of Organic Agriculture (FiBL), Frick, International Federation of Organic Agriculture Movements (IFOAM), Bonn. |
| [13] |
Letourneau DK, Bothwell SG (2008) Comparison of organic and conventional farms: Challenging ecologists to make biodiversity functional. Front Ecol Environ 6: 430–438. doi: 10.1890/070081
|
| [14] |
Wolfe MS, Baresel JP, Desclaux D, et al. (2008) Developments in breeding cereals for organic agriculture. Euphytica 163: 323–346. doi: 10.1007/s10681-008-9690-9
|
| [15] |
Lotter DW (2003) Organic agriculture. J Sustain Agr 21: 59–128. doi: 10.1300/J064v21n04_06
|
| [16] |
Watson CA, Atkinson D, Gosling P, et al. (2002) Managing soil fertility in organic farming systems. Soil Use Manage 18: 239–247. doi: 10.1079/SUM2002131
|
| [17] |
Gosling P, Ozaki A, Jones J, et al. (2010) Organic management of tilled agricultural soils results in a rapid increase in colonisation potential and spore populations of arbuscular mycorrhizal fungi. Agr Ecosyst Environ 139: 273–279. doi: 10.1016/j.agee.2010.08.013
|
| [18] |
Bengtsson J, Ahnström J, Weibull AC (2005) The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J Appl Ecol 42: 261–269. doi: 10.1111/j.1365-2664.2005.01005.x
|
| [19] | Costanzo A, Bàrberi P (2013) Functional agrobiodiversity and agroecosystem services in sustainable wheat production. A review. Agron Sustain Dev 34: 327–348. |
| [20] |
Gosling P, Shepherd M (2005) Long-term changes in soil fertility in organic arable farming systems in England, with particular reference to phosphorus and potassium. Agr Ecosyst Environ 105: 425–432. doi: 10.1016/j.agee.2004.03.007
|
| [21] |
Lenzi A, Antichi D, Bigongiali F, et al. (2009) Effect of different cover crops on organic tomato production. Renew Agr Food Syst 24: 92–101. doi: 10.1017/S1742170508002445
|
| [22] | Jeffries P, Gianinazzi S, Turnau K, et al. (2003) The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol Fert Soils 37: 1–16. |
| [23] |
Gianinazzi S, Gollotte A, Binet MN, et al. (2010) Agroecology: the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 20: 519–530. doi: 10.1007/s00572-010-0333-3
|
| [24] | Smith SE, Read D (2008) Mycorrhizal symbiosis (Third Edition), Academic Press, London. |
| [25] | Berta G, Copetta A, Gamalero E, et al. (2013) Maize development and grain quality are differentially affected by mycorrhizal fungi and a growth-promoting pseudomonad in the field. Mycorrhiza 24: 161–170. |
| [26] |
Rinaudo V, Barberi P, Giovannetti M, et al. (2010) Mycorrhizal fungi suppress aggressive agricultural weeds. Plant Soil 333: 7–20. doi: 10.1007/s11104-009-0202-z
|
| [27] |
Veiga RSL, Jansa J, Frossard E, et al. (2011) Can Arbuscular mycorrhizal fungi reduce the growth of agricultural weeds? PLoS One 6: e27825. doi: 10.1371/journal.pone.0027825
|
| [28] |
Veresoglou SD, Rillig MC (2012) Suppression of fungal and nematode plant pathogens through arbuscular mycorrhizal fungi. Biol Letters 8: 214–217. doi: 10.1098/rsbl.2011.0874
|
| [29] |
Augé RM (2001) Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 11: 3–42. doi: 10.1007/s005720100097
|
| [30] | Pavithra D, Yapa N (2018) Arbuscular mycorrhizal fungi inoculation enhances drought stress tolerance of plants. Groundwater for Sustainable Development. |
| [31] | Bothe H, Regvar M, Turnau K (2010) Arbuscular mycorrhiza, heavy metal, and salt tolerance, soil heavy metals, Springer Berlin Heidelberg. 87–111. |
| [32] |
Forgy D (2012) Arbuscular mycorrhizal fungi can benefit heavy metal tolerance and phytoremediation. J Nat Resour Life Sci Educ 41: 23–26. doi: 10.4195/jnrlse.2012.0001se
|
| [33] |
Wang Y, Wang M, Li Y, et al. (2018) Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS One 13: e0196408. doi: 10.1371/journal.pone.0196408
|
| [34] | Miransari M (2017) Arbuscular mycorrhizal fungi and soil salinity, in: Johnson, N.C. et al. Eds., Mycorrhizal Mediation of Soil, Elsevier, 263–277. |
| [35] |
Mathur S, Sharma MP, Jajoo A (2018) Improved photosynthetic efficacy of maize (Zea mays) plants with arbuscular mycorrhizal fungi (AMF) under high temperature stress. J Photoch Photobio B 180: 149–154. doi: 10.1016/j.jphotobiol.2018.02.002
|
| [36] | Zhu X, Song F, Liu F (2017) Arbuscular mycorrhizal fungi and tolerance of temperature stress in plants, in: Wu, Q.S. Eds., Arbuscular Mycorrhizas and Stress Tolerance of Plants, Springer Singapore, Singapore, 163–194. |
| [37] |
Rillig MC (2004) Arbuscular mycorrhizae, glomalin, and soil aggregation. Can J Soil Sci 84: 355–363. doi: 10.4141/S04-003
|
| [38] |
Bedini S, Pellegrino E, Avio L, et al. (2009) Changes in soil aggregation and glomalin-related soil protein content as affected by the arbuscular mycorrhizal fungal species Glomus mosseae and Glomus intraradices. Soil Biol Biochem 41: 1491–1496. doi: 10.1016/j.soilbio.2009.04.005
|
| [39] |
Cardoso IM, Kuyper TW (2006) Mycorrhizas and tropical soil fertility. Agr Ecosyst Environ 116: 72–84. doi: 10.1016/j.agee.2006.03.011
|
| [40] | Giovannetti M, Avio L (2002) Biotechnology of arbuscular mycorrhizas, in: GK George and KA Dilip, Eds., Applied Mycology and Biotechnology, Elsevier, 275–310. |
| [41] |
Wright SF, Upadhyaya A (1996) Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci 161: 575–586. doi: 10.1097/00010694-199609000-00003
|
| [42] | Njeru EM, Avio L, Sbrana C, et al. (2013) First evidence for a major cover crop effect on arbuscular mycorrhizal fungi and organic maize growth. Agron Sustain Dev 34: 841–848. |
| [43] |
Avio L, Castaldini M, Fabiani A, et al. (2013) Impact of nitrogen fertilization and soil tillage on arbuscular mycorrhizal fungal communities in a Mediterranean agroecosystem. Soil Biol Biochem 67: 285–294. doi: 10.1016/j.soilbio.2013.09.005
|
| [44] |
Oehl F, Sieverding E, Mäder P, et al. (2004) Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 138: 574–583. doi: 10.1007/s00442-003-1458-2
|
| [45] |
Mäder P, Edenhofer S, Boller T, et al. (2000) Arbuscular mycorrhizae in a long-term field trial comparing low-input (organic, biological) and high-input (conventional) farming systems in a crop rotation. Biol Fert Soils 31: 150–156. doi: 10.1007/s003740050638
|
| [46] | Njeru EM, Avio L, Bocci G, et al. (2014) Contrasting effects of cover crops on 'hot spot' arbuscular mycorrhizal fungal communities in organic tomato. Biol Fert Soils 51: 151–166. |
| [47] |
Raviv M (2010) The use of mycorrhiza in organically-grown crops under semi arid conditions: A review of benefits, constraints and future challenges. Symbiosis 52: 65–74. doi: 10.1007/s13199-010-0089-8
|
| [48] |
Karasawa T, Takebe M (2012) Temporal or spatial arrangements of cover crops to promote arbuscular mycorrhizal colonization and P uptake of upland crops grown after nonmycorrhizal crops. Plant Soil 353: 355–366. doi: 10.1007/s11104-011-1036-z
|
| [49] |
Lehmann A, Barto EK, Powell J, et al. (2012) Mycorrhizal responsiveness trends in annual crop plants and their wild relatives-a meta-analysis on studies from 1981 to 2010. Plant Soil 355: 231–250. doi: 10.1007/s11104-011-1095-1
|
| [50] |
Conversa G, Lazzizera C, Bonasia A, et al. (2013) Yield and phosphorus uptake of a processing tomato crop grown at different phosphorus levels in a calcareous soil as affected by mycorrhizal inoculation under field conditions. Biol Fert Soils 49: 691–703. doi: 10.1007/s00374-012-0757-3
|
| [51] |
Douds Jr DD, Reider C (2003) Inoculation with mycorrhizal fungi increases the yield of green peppers in a high P soil. Biol Agric Hortic 21: 91–102. doi: 10.1080/01448765.2003.9755251
|
| [52] |
Bedini S, Avio L, Sbrana C, et al. (2013) Mycorrhizal activity and diversity in a long-term organic Mediterranean agroecosystem. Biol Fert Soils 49: 781–790. doi: 10.1007/s00374-012-0770-6
|
| [53] |
Harrier LA, Watson CA (2004) The potential role of arbuscular mycorrhizal (AM) fungi in the bioprotection of plants against soil-borne pathogens in organic and/or other sustainable farming systems. Pest Manag Sci 60: 149–157. doi: 10.1002/ps.820
|
| [54] | Ryan MH, Tibbett M (2008) The role of arbuscular mycorrhizas in organic farming, in: Kirchmann, H. Bergström, L. Eds., Organic Crop Production-Ambitions and Limitations Springer Netherlands. 189–229. |
| [55] |
Verbruggen E, Röling WFM, Gamper HA, et al. (2010) Positive effects of organic farming on below-ground mutualists: Large-scale comparison of mycorrhizal fungal communities in agricultural soils. New Phytol 186: 968–979. doi: 10.1111/j.1469-8137.2010.03230.x
|
| [56] |
Van der Heijden MGA, Klironomos JN, Ursic M, et al. (1998) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396: 69–72. doi: 10.1038/23932
|
| [57] |
Wang F, Hu J, Lin X, et al. (2011) Arbuscular mycorrhizal fungal community structure and diversity in response to long-term fertilization: A field case from China. World J Microbiol Biotechnol 27: 67–74. doi: 10.1007/s11274-010-0427-2
|
| [58] |
Hooper DU, Chapin FS, Ewel JJ, et al. (2005) Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol Monogr 75: 3–35. doi: 10.1890/04-0922
|
| [59] |
Fargione J, Tilman D, Dybzinski R, et al. (2007) From selection to complementarity: Shifts in the causes of biodiversity–productivity relationships in a long-term biodiversity experiment. Proc R Soc B: Biol Sci 274: 871–876. doi: 10.1098/rspb.2006.0351
|
| [60] |
Gosling P, Hodge A, Goodlass G, et al. (2006) Arbuscular mycorrhizal fungi and organic farming. Agr Ecosyst Environ 113: 17–35. doi: 10.1016/j.agee.2005.09.009
|
| [61] |
Njeru EM, Bocci G, Avio L, et al. (2017) Functional identity has a stronger effect than diversity on mycorrhizal symbiosis and productivity of field grown organic tomato. Eur J Agron 86: 1–11. doi: 10.1016/j.eja.2017.02.007
|
| [62] |
Costanzo A, Bàrberi P (2016) Field scale functional agrobiodiversity in organic wheat: Effects on weed reduction, disease susceptibility and yield. Eur J Agron 76: 1–16. doi: 10.1016/j.eja.2016.01.012
|
| [63] |
Gomiero T, Pimentel D, Paoletti MG (2011) Environmental impact of different agricultural management practices: Conventional vs. organic agriculture. Crit Rev Plant Sci 30: 95–124. doi: 10.1080/07352689.2011.554355
|
| [64] |
Kabir Z, Koide RT (2002) Effect of autumn and winter mycorrhizal cover crops on soil properties, nutrient uptake and yield of sweet corn in Pennsylvania, USA. Plant Soil 238: 205–215. doi: 10.1023/A:1014408723664
|
| [65] |
Weil R, Kremen A (2007) Thinking across and beyond disciplines to make cover crops pay. J Sci Food Agric 87: 551–557. doi: 10.1002/jsfa.2742
|
| [66] |
Bàrberi P, Mazzoncini M (2001) Changes in weed community composition as influenced by cover crop and management system in continuous corn. Weed Sci 49: 491–499. doi: 10.1614/0043-1745(2001)049[0491:CIWCCA]2.0.CO;2
|
| [67] | Creamer NG, Bennett MA, Stinner BR, et al. (1996) A comparison of four processing tomato production systems differing in cover crop and chemical inputs. J Am Soc Hortic Sci 121: 559–568. |
| [68] |
Lawson A, Fortuna AM, Cogger C, et al. (2013) Nitrogen contribution of rye–hairy vetch cover crop mixtures to organically grown sweet corn. Renew Agr Food Syst 28: 59–69. doi: 10.1017/S1742170512000014
|
| [69] |
Carrera LM, Buyer JS, Vinyard B, et al. (2007) Effects of cover crops, compost, and manure amendments on soil microbial community structure in tomato production systems. Appl Soil Ecol 37: 247–255. doi: 10.1016/j.apsoil.2007.08.003
|
| [70] |
Buyer JS, Teasdale JR, Roberts DP, et al. (2010) Factors affecting soil microbial community structure in tomato cropping systems. Soil Biol Biochem 42: 831–841. doi: 10.1016/j.soilbio.2010.01.020
|
| [71] |
Boswell EP, Koide RT, Shumway DL, et al (1998) Winter wheat cover cropping, VA mycorrhizal fungi and maize growth and yield. Agr Ecosyst Environ 67: 55–65. doi: 10.1016/S0167-8809(97)00094-7
|
| [72] |
Kabir Z, Koide RT (2000) The effect of dandelion or a cover crop on mycorrhiza inoculum potential, soil aggregation and yield of maize. Agr Ecosyst Environ 78: 167–174. doi: 10.1016/S0167-8809(99)00121-8
|
| [73] |
Lehman RM, Taheri WI, Osborne SL, et al. (2012) Fall cover cropping can increase arbuscular mycorrhizae in soils supporting intensive agricultural production. Appl Soil Ecol 61: 300–304. doi: 10.1016/j.apsoil.2011.11.008
|
| [74] |
Galvez L, Douds DD, Wagoner P, et al. (1995) An overwintering cover crop increases inoculum of VAM fungi in agricultural soil. Am J Altern Agric 10: 152–156. doi: 10.1017/S0889189300006391
|
| [75] | Black R, Tinker PB (1979) The development of endomycorrhizal root systems II. Effect of agronomic factors and soil conditions on the development of vesicular-arbuscular mycorrhizal infection in barley and on the endophyte spore density. New Phytol 83: 401–413. |
| [76] |
Monreal MA, Grant CA, Irvine RB, et al. (2011) Crop management effect on arbuscular mycorrhizae and root growth of flax. Can J Plant Sci 91: 315–324. doi: 10.4141/CJPS10136
|
| [77] |
Gavito ME, Miller MH (1998) Changes in mycorrhiza development in maize induced by crop management practices. Plant Soil 198: 185–192. doi: 10.1023/A:1004314406653
|
| [78] |
White CM, Weil RR (2010) Forage radish and cereal rye cover crop effects on mycorrhizal fungus colonization of maize roots. Plant Soil 328: 507–521. doi: 10.1007/s11104-009-0131-x
|
| [79] | Pellerin S, Mollier A, Morel C, et al. (2007) Effect of incorporation of Brassica napus L. residues in soils on mycorrhizal fungus colonisation of roots and phosphorus uptake by maize (Zea mays L.). Eur J Agron 26: 113–120. |
| [80] |
Moonen AC, Bàrberi P (2006) An ecological approach to study the physical and chemical effects of rye cover crop residues on Amaranthus retroflexus, Echinochloa crus-galli and maize. Ann Appl Biol 148: 73–89. doi: 10.1111/j.1744-7348.2006.00038.x
|
| [81] |
Wyland LJ, Jackson LE, Chaney WE, et al. (1996) Winter cover crops in a vegetable cropping system: Impacts on nitrate leaching, soil water, crop yield, pests and management costs. Agr Ecosyst Environ 59: 1–17. doi: 10.1016/0167-8809(96)01048-1
|
| [82] | Creamer NG, Bennett MA, Stinner BR (1997) Evaluation of cover crop mixtures for use in vegetable production systems. Hort Sci 32: 866–870. |
| [83] |
Groff S (2008) Mixtures and cocktails: Soil is meant to be covered. J Soil Water Conserv 63: 110–111. doi: 10.2489/jswc.63.4.110A
|
| [84] |
Koide RT, Peoples MS (2012) On the nature of temporary yield loss in maize following canola. Plant Soil 360: 259–269. doi: 10.1007/s11104-012-1237-0
|
| [85] |
Karasawa T, Kasahara Y, Takebe M (2001) Variable response of growth and arbuscular mycorrhizal colonization of maize plants to preceding crops in various types of soils. Biol Fert Soils 33: 286–293. doi: 10.1007/s003740000321
|
| [86] | Antunes PM, Goss MJ (2005) Communication in the tripartite symbiosis formed by arbuscular mycorrhizal fungi, rhizobia and legume plants: A review, Agronomy Monograph No.48, Madison: American Society of Agronomy, 199–222. |
| [87] |
de Varennes A, Goss MJ (2007) The tripartite symbiosis between legumes, rhizobia and indigenous mycorrhizal fungi is more efficient in undisturbed soil. Soil Biol Biochem 39: 2603–2607. doi: 10.1016/j.soilbio.2007.05.007
|
| [88] |
Oruru MB, Njeru EM, Pasquet R, et al. (2018) Response of a wild-type and modern cowpea cultivars to arbuscular mycorrhizal inoculation in sterilized and non-sterilized soil. J Plant Nutr 41: 90–101. doi: 10.1080/01904167.2017.1381728
|
| [89] | Verma M, Brar SK, Tyagi RD, et al. (2007) Antagonistic fungi, Trichoderma spp.: Panoply of biological control. Biochem Eng J 37: 1–20. |
| [90] |
Vatovec C, Jordan N, Huerd S (2005) Responsiveness of certain agronomic weed species to arbuscular mycorrhizal fungi. Renew Agr Food Syst 20: 181–189. doi: 10.1079/RAF2005115
|
| [91] | Atkinson D, Baddeley JA, Goicoechea N, et al. (2002) Arbuscular mycorrhizal fungi in low input agriculture, ycorrhizal technology in agriculture, 211–222. |
| [92] |
Nyamwange MM, Njeru EM, Mucheru-Muna M, et al. (2018) Soil management practices affect arbuscular mycorrhizal fungi propagules, root colonization and growth of rainfed maize. AIMS Agric Food 3: 120–134. doi: 10.3934/agrfood.2018.2.120
|
| [93] |
Lammerts van Bueren ET, Jones SS, Tamm L, et al. (2011) The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS Wagen J Life Sci 58: 193–205. doi: 10.1016/j.njas.2010.04.001
|
| [94] |
Zhu YG, Smith SE, Barritt AR, et al. (2001) Phosphorus (P) efficiencies and mycorrhizal responsiveness of old and modern wheat cultivars. Plant Soil 237: 249–255. doi: 10.1023/A:1013343811110
|
| [95] |
Singh AK, Hamel C, DePauw RM, et al. (2012) Genetic variability in arbuscular mycorrhizal fungi compatibility supports the selection of durum wheat genotypes for enhancing soil ecological services and cropping systems in Canada. Can J Microbiol 58: 293–302. doi: 10.1139/w11-140
|
| [96] |
Tawaraya K (2003) Arbuscular mycorrhizal dependency of different plant species and cultivars. Soil Sci Plant Nutr 49: 655–668. doi: 10.1080/00380768.2003.10410323
|
| [97] |
Döring TF, Knapp S, Kovacs G, et al. (2011) Evolutionary Plant Breeding in Cereals-Into a New Era. Sustainability 3: 1944–1971. doi: 10.3390/su3101944
|
| [98] |
Kiær LP, Skovgaard IM, Østergard H (2009) Grain yield increase in cereal variety mixtures: A meta-analysis of field trials. Field Crops Res 114: 361–373. doi: 10.1016/j.fcr.2009.09.006
|
| [99] | Oruru MB, Njeru EM (2016) Upscaling arbuscular mycorrhizal symbiosis and related agroecosystems services in smallholder farming systems. Biomed Res Int 2016: 4376240. |
| [100] |
Schwartz MW, Hoeksema JD, Gehring CA, et al. (2006) The promise and the potential consequences of the global transport of mycorrhizal fungal inoculum. Ecol Lett 9: 501–515. doi: 10.1111/j.1461-0248.2006.00910.x
|
| [101] |
Wang FY, Tong RJ, Shi ZY, et al. (2011) Inoculations with arbuscular mycorrhizal fungi increase vegetable yields and decrease phoxim concentrations in carrot and green onion and their soils. PLoS One 6: e16949. doi: 10.1371/journal.pone.0016949
|
| [102] |
Douds Jr DD, Nagahashi G, Reider C, et al. (2007) Inoculation with arbuscular mycorrhizal fungi increases the yield of potatoes in a high P soil. Biol Agric Hortic 25: 67–78. doi: 10.1080/01448765.2007.10823209
|
| [103] | Muok BO, Matsumura A, Ishii T, et al (2009) The effect of intercropping Sclerocarya birrea (A. Rich.) Hochst, millet and corn in the presence of arbuscular mycorrhizal fungi. Afr J Biotechnol 8: 807–812. |
| [104] |
White JA, Tallaksen J, Charvat I (2008) The effects of arbuscular mycorrhizal fungal inoculation at a roadside prairie restoration site. Mycologia 100: 6–11. doi: 10.1080/15572536.2008.11832494
|
| [105] | Garland BC, Schroeder-Moreno MS, Fernandez GE, et al (2011) Influence of summer cover crops and mycorrhizal fungi on strawberry production in the Southeastern United States. HortSci 46: 985–991. |
| [106] |
Douds Jr DD, Millner PD (1999) Biodiversity of arbuscular mycorrhizal fungi in agroecosystems. Agr Ecosyst Environ 74: 77–93. doi: 10.1016/S0167-8809(99)00031-6
|
| [107] |
Saito M, Marumoto T (2002) Inoculation with arbuscular mycorrhizal fungi: The status quo in Japan and the future prospects. Plant Soil 244: 273–279. doi: 10.1023/A:1020287900415
|
| [108] | Brito I, De Carvalho M, Goss MJ (2011) Summer survival of arbuscular mycorrhiza extraradical mycelium and the potential for its management through tillage options in Mediterranean cropping systems. Soil Use Manage 27: 350–356. |
| [109] |
Antunes P, Koch A, Dunfield K, et al. (2009) Influence of commercial inoculation with Glomus intraradices on the structure and functioning of an AM fungal community from an agricultural site. Plant Soil 317: 257–266. doi: 10.1007/s11104-008-9806-y
|
| [110] | Campanelli A, Ruta C, Tagarelli A, et al. (2011) Nursery inoculation with the arbuscular mycorrhizal fungus Glomus viscosum and its effect on the growth and physiology of hybrid artichoke seedlings. Ital J Agron 6: 159–164. |
| [111] | An GH, Kobayashi S, Enoki H, et al. (2010) How does arbuscular mycorrhizal colonization vary with host plant genotype? An example based on maize germplasms. Plant Soil 327: 441–453. |
| [112] | Turrini A, Giordani T, Avio L, et al. (2015) Large variation in mycorrhizal colonization among wild accessions, cultivars, and inbreds of sunflower (Helianthus annuus L.). Euphytica 207: 331–342. |
| [113] |
Pellegrino E, Turrini A, Gamper HA, et al. (2012) Establishment, persistence and effectiveness of arbuscular mycorrhizal fungal inoculants in the field revealed using molecular genetic tracing and measurement of yield components. New Phytol 194: 810–822. doi: 10.1111/j.1469-8137.2012.04090.x
|
| [114] |
Janoušková M, Krak K, Wagg C, et al (2013) Effects of inocula additions in presence of a pre-established arbuscular mycorrhizal fungal community. Appl Environ Microbiol 79: 6507– 6515. doi: 10.1128/AEM.02135-13
|
| [115] |
Brussaard L, de Ruiter PC, Brown GG (2007) Soil biodiversity for agricultural sustainability. Agr Ecosyst Environ 121: 233–244. doi: 10.1016/j.agee.2006.12.013
|
| [116] |
Tooker JF, Frank SD (2012) Genotypically diverse cultivar mixtures for insect pest management and increased crop yields. J Appl Ecol 49: 974–985. doi: 10.1111/j.1365-2664.2012.02173.x
|
| [117] |
Himanen SJ, Ketoja E, Hakala K, et al. (2013) Cultivar diversity has great potential to increase yield of feed barley. Agron Sustain Dev 33: 519–530. doi: 10.1007/s13593-012-0120-y
|
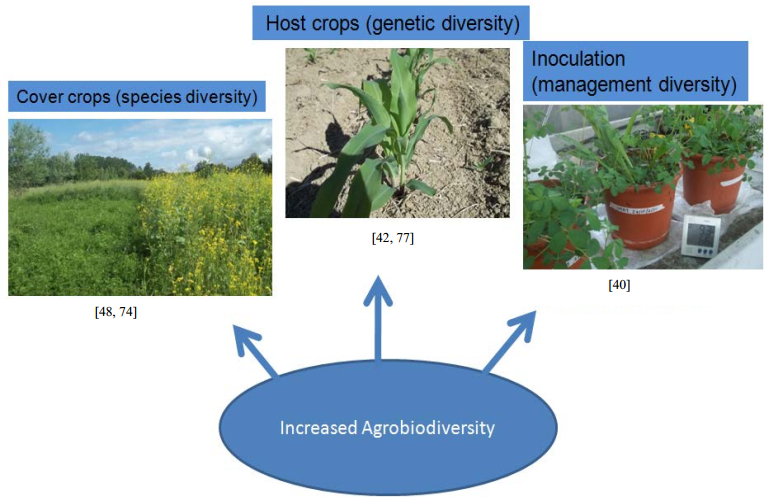
Figures(1)

Ezekiel Mugendi Njeru. Exploiting diversity to promote arbuscular mycorrhizal symbiosis and crop productivity in organic farming systems[J]. AIMS Agriculture and Food, 2018, 3(3): 280-294. doi: 10.3934/agrfood.2018.3.280







 DownLoad:
DownLoad: