Citation: Rosa Molfetta, Beatrice Zitti, Angela Santoni, Rossella Paolini. Ubiquitin and ubiquitin-like modifiers modulate NK cell-mediated recognition and killing of damaged cells[J]. AIMS Allergy and Immunology, 2017, 1(4): 164-180. doi: 10.3934/Allergy.2017.4.164

| [1] |
Caligiuri MA (2008) Human natural killer cells. Blood 112: 461–469. doi: 10.1182/blood-2007-09-077438

|
| [2] | Morvan MG, Lanier LL (2016) NK cells and cancer: you can teach innate cells new tricks. Nat Rev Cancer 16: 7–19. |
| [3] |
Long EO, Kim HS, Liu D, et al. (2013) Controlling natural killer cell responses: integration of signals for activation and inhibition. Annu Rev Immunol 31: 227–258. doi: 10.1146/annurev-immunol-020711-075005
|
| [4] |
Raulet DH, Gasser S, Gowen BG, et al. (2013) Regulation of ligands for the NKG2D activating receptor. Annu Rev Immunol 31: 413–441. doi: 10.1146/annurev-immunol-032712-095951
|
| [5] | Cerboni C, Fionda C, Soriani A, et al. (2014) The DNA damage response: a common pathway in the regulation of NKG2D and DNAM-1 ligand expression in normal, infected, and cancer cells. Front Immunol 4: 508. |
| [6] |
Chan CJ, Smyth MJ, Martinet L (2014) Molecular mechanisms of natural killer cell activation in response to cellular stress. Cell Death Differ 21: 5–14. doi: 10.1038/cdd.2013.26
|
| [7] |
Ullrich E, Koch J, Cerwenka A, et al. (2013) New prospects on the NKG2D/NKG2DL system for oncology. Oncoimmunology 2: e26097. doi: 10.4161/onci.26097
|
| [8] |
Marcus A, Gowen BG, Thompson TW, et al. (2014) Recognition of tumors by the innate immune system and natural killer cells. Adv Immunol 122: 91–128. doi: 10.1016/B978-0-12-800267-4.00003-1
|
| [9] |
Lanier LL (2015) NKG2D receptor and its ligands in host defense. Cancer Immunol Res 3: 575–582. doi: 10.1158/2326-6066.CIR-15-0098
|
| [10] |
Guerra N, Joncker NT, Choy A, et al. (2008) NKG2D-deficient mice are defective in tumor surveillance in models of spontaneous malignancy. Immunity 28: 571–580. doi: 10.1016/j.immuni.2008.02.016
|
| [11] |
Zafirova B, Mandarić S, Antulov R, et al. (2009) Altered NK cell development and enhanced NK cell-mediated resistance to mouse cytomegalovirus in NKG2D-deficient mice. Immunity 31: 270–282. doi: 10.1016/j.immuni.2009.06.017
|
| [12] |
Bauer S, Groh V, Wu J, et al. (1999) Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285: 727–729. doi: 10.1126/science.285.5428.727
|
| [13] |
Wu J, Song Y, Bakker AB, et al. (1999) An activating immunoreceptor complex formed by NKG2D and DAP10. Science 285: 730–732. doi: 10.1126/science.285.5428.730
|
| [14] |
Upshaw JL, Leibson PJ (2006) NKG2D-mediated activation of cytotoxic lymphocytes: unique signaling pathways and distinct functional outcomes. Semin Immunol 18: 167–175. doi: 10.1016/j.smim.2006.03.001
|
| [15] |
Diefenbach A, Tomasello E, Lucas M, et al. (2002) Selective associations with signaling proteins determine stimulatory versus costimulatory activity of NKG2D. Nat Immunol 3: 1142–1149. doi: 10.1038/ni858
|
| [16] |
Gilfillan S, Ho EL, Cella M, et al. (2002) NKG2D recruits two distinct adapters to trigger NK cell activation and costimulation. Nat Immunol 3: 1150–1155. doi: 10.1038/ni857
|
| [17] |
Eagle RA, Trowsdale J (2007) Promiscuity and the single receptor: NKG2D. Nat Rev Immunol 7: 737–744. doi: 10.1038/nri2144
|
| [18] | de Andrade LF, Smyth MJ, Martinet L (2013) DNAM-1 control of natural killer cells functions through nectin and nectin-like proteins. Immunol Cell Biol 92: 237–244. |
| [19] |
Reymond N, Imbert AM, Devilard E, et al. (2004) DNAM-1 and PVR regulate monocyte migration through endothelial junctions. J Exp Med 199: 1331–1341. doi: 10.1084/jem.20032206
|
| [20] |
Iguchi-Manaka A, Kai H, Yamashita Y, et al. (2008) Accelerated tumor growth in mice deficient in DNAM-1 receptor. J Exp Med 205: 2959–2964. doi: 10.1084/jem.20081611
|
| [21] |
Gilfillan S, Chan CJ, Cella M, et al. (2008) DNAM-1 promotes activation of cytotoxic lymphocytes by nonprofessional antigen-presenting cells and tumors. J Exp Med 205: 2965–2973. doi: 10.1084/jem.20081752
|
| [22] |
Welch MJ, Teijaro JR, Lewicki HA, et al. (2012) CD8 T cell defect of TNF-α and IL-2 in DNAM-1 deficient mice delays clearance in vivo of a persistent virus infection. Virology 429: 163–170. doi: 10.1016/j.virol.2012.04.006
|
| [23] | Shibuya A, Lanier LL, Phillips JH (1998) Protein kinase C is involved in the regulation of both signaling and adhesion mediated by DNAX accessory molecule-1 receptor. J Immunol 161: 1671–1676. |
| [24] |
Shibuya K, Lanier LL, Phillips JH, et al. (1999) Physical and functional association of LFA-1 with DNAM-1 adhesion molecule. Immunity 11: 615–623. doi: 10.1016/S1074-7613(00)80136-3
|
| [25] |
Bottino C, Castriconi R, Pende D, et al. (2003) Identification of PVR (CD155) and Nectin-2 (CD112) as cell surface ligands for the human DNAM-1 (CD226) activating molecule. J Exp Med 198: 557–567. doi: 10.1084/jem.20030788
|
| [26] |
Tahara-Hanaoka S, Shibuya K, Onoda Y, et al. (2004) Functional characterization of DNAM-1 (CD226) interaction with its ligands PVR (CD155) and nectin-2 (PRR-2/CD112). Int Immunol 16: 533–538. doi: 10.1093/intimm/dxh059
|
| [27] |
Fuchs A, Colonna M (2006) The role of NK cell recognition of nectin and nectin-like proteins in tumor immunosurveillance. Semin Cancer Biol 16: 359–366. doi: 10.1016/j.semcancer.2006.07.002
|
| [28] |
Stanietsky N, Rovis TL, Glasner A, et al. (2013) Mouse TIGIT inhibits NK-cell cytotoxicity upon interaction with PVR. Eur J Immunol 43: 2138–2150. doi: 10.1002/eji.201243072
|
| [29] |
Pende D, Cantoni C, Rivera P, et al. (2001) Role of NKG2D in tumor cell lysis mediated by human NK cells: cooperation with natural cytotoxicity receptors and capability of recognizing tumors of nonepithelial origin. Eur J Immunol 31: 1076–1086. doi: 10.1002/1521-4141(200104)31:4<1076::AID-IMMU1076>3.0.CO;2-Y
|
| [30] |
Jinushi M, Takehara T, Tatsumi T, et al. (2003) Expression and role of MICA and MICB in human hepatocellular carcinomas and their regulation by retinoic acid. Int J Cancer 104: 354–361. doi: 10.1002/ijc.10966
|
| [31] |
Salih HR, Antropius H, Gieseke F, et al. (2003) Functional expression and release of ligands for the activating immunoreceptor NKG2D in leukemia. Blood 102: 1389–1396. doi: 10.1182/blood-2003-01-0019
|
| [32] | Friese MA, Platten M, Lutz SZ, et al. (2003) MICA/NKG2D-mediated immunogene therapy of experimental gliomas. Cancer Res 63: 8996–9006. |
| [33] |
Groh V, Rhinehart R, Secrist H, et al. (1999) Broad tumor-associated expression and recognition by tumor-derived gamma delta T cells of MICA and MICB. Proc Natl Acad Sci USA 96: 6879–6884. doi: 10.1073/pnas.96.12.6879
|
| [34] |
Pende D, Spaggiari GM, Marcenaro S, et al. (2005) Analysis of the receptor-ligand interactions in the natural killer-mediated lysis of freshly isolated myeloid or lymphoblastic leukemias: evidence for the involvement of the Poliovirus receptor (CD155) and Nectin-2 (CD112). Blood 105: 2066–2073. doi: 10.1182/blood-2004-09-3548
|
| [35] |
Carlsten M, Björkström NK, Norell H, et al. (2007) DNAX accessory molecule-1 mediated recognition of freshly isolated ovarian carcinoma by resting natural killer cells. Cancer Res 67: 1317–1325. doi: 10.1158/0008-5472.CAN-06-2264
|
| [36] |
El-Sherbiny YM, Meade JL, Holmes TD, et al. (2007) The requirement for DNAM-1, NKG2D, and NKp46 in the natural killer cell-mediated killing of myeloma cells. Cancer Res 67: 8444–8449. doi: 10.1158/0008-5472.CAN-06-4230
|
| [37] |
Lakshmikanth T, Burke S, Ali TH, et al. (2009) NCRs and DNAM-1 mediate NK cell recognition and lysis of human and mouse melanoma cell lines in vitro and in vivo. J Clin Invest 119: 1251–1263. doi: 10.1172/JCI36022
|
| [38] |
Torelli GF, Peragine N, Raponi S, et al. (2014) Recognition of adult and pediatric acute lymphoblastic leukemia blasts by natural killer cells. Haematologica 99: 1248–1254. doi: 10.3324/haematol.2013.101931
|
| [39] |
Chan CJ, Andrews DM, McLaughlin NM, et al. (2010) DNAM-1/CD155 interactions promote cytokine and NK cell-mediated suppression of poorly immunogenic melanoma metastases. J Immunol 184: 902–911. doi: 10.4049/jimmunol.0903225
|
| [40] | Paolini R, Bernardini G, Molfetta R, et al. (2015) NK cells and interferons. Cytokine Growth F R 2: 113–120. |
| [41] |
Groh V, Rhinehart R, Randolph-Habecker J, et al. (2001) Costimulation of CD8alphabeta T cells by NKG2D via engagement by MIC induced on virus-infected cells. Nat Immunol 2: 255–260. doi: 10.1038/85321
|
| [42] |
Cosman D, Müllberg J, Sutherland CL, et al. (2001) ULBPs, novel MHC class I-related molecules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity through the NKG2D receptor. Immunity 14: 123–133. doi: 10.1016/S1074-7613(01)00095-4
|
| [43] |
Welte SA, Sinzger C, Lutz SZ, et al. (2003) Selective intracellular retention of virally induced NKG2D ligands by the human cytomegalovirus UL16 glycoprotein. Eur J Immunol 33: 194–203. doi: 10.1002/immu.200390022
|
| [44] |
Rölle A, Mousavi-Jazi M, Eriksson M, et al. (2003) Effects of human cytomegalovirus infection on ligands for the activating NKG2D receptor of NK cells: up-regulation of UL16-binding protein (ULBP)1 and ULBP2 is counteracted by the viral UL16 protein. J Immunol 171: 902–908. doi: 10.4049/jimmunol.171.2.902
|
| [45] | Tomasec P, Wang EC, Davison AJ, et al. (2005) Downregulation of natural killer cell-activating ligand CD155 by human cytomegalovirus UL141. Nat Immunol 6: 181–188. |
| [46] |
Cerboni C, Neri F, Casartelli N, et al. (2007) Human immunodeficiency virus 1 Nef protein downmodulates the ligands of the activating receptor NKG2D and inhibits natural killer cell-mediated cytotoxicity. J Gen Virol 88: 242–250. doi: 10.1099/vir.0.82125-0
|
| [47] |
Ward J, Bonaparte M, Sacks J, et al. (2007) HIV modulates the expression of ligands important in triggering natural killer cell cytotoxic responses on infected primary T-cell blasts. Blood 110: 1207–1214. doi: 10.1182/blood-2006-06-028175
|
| [48] |
Matusali G, Potestà M, Santoni A, et al. (2012) The human immunodeficiency virus type 1 Nef and Vpu proteins downregulate the natural killer cell-activating ligand PVR. J Virol 86: 4496–4504. doi: 10.1128/JVI.05788-11
|
| [49] |
Pappworth IY, Wang EC, Rowe M (2007) The switch from latent to productive infection in epstein-barr virus-infected B cells is associated with sensitization to NK cell killing. J Virol 81: 474–482. doi: 10.1128/JVI.01777-06
|
| [50] |
Sirén J, Sareneva T, Pirhonen J, et al. (2004) Cytokine and contact-dependent activation of natural killer cells by influenza A or Sendai virus-infected macrophages. J Gen Virol 85: 2357–2364. doi: 10.1099/vir.0.80105-0
|
| [51] |
Cerboni C, Zingoni A, Cippitelli M, et al. (2007) Antigen-activated human T lymphocytes express cell-surface NKG2D ligands via an ATM/ATR-dependent mechanism and become susceptible to autologous NK-cell lysis. Blood 110: 606–615. doi: 10.1182/blood-2006-10-052720
|
| [52] |
Ardolino M, Zingoni A, Cerboni C, et al. (2011) DNAM-1 ligand expression on Ag-stimulated T lymphocytes is mediated by ROS-dependent activation of DNA-damage response: relevance for NK-T cell interaction. Blood 117: 4778–4786. doi: 10.1182/blood-2010-08-300954
|
| [53] |
Ward J, Davis Z, DeHart J, et al. (2009) HIV-1 Vpr triggers natural killer cell-mediated lysis of infected cells through activation of the ATR-mediated DNA damage response. PLoS Pathog 5: e1000613. doi: 10.1371/journal.ppat.1000613
|
| [54] |
Richard J, Sindhu S, Pham TN, et al. (2010) HIV-1 Vpr up-regulates expression of ligands for the activating NKG2D receptor and promotes NK cell-mediated killing. Blood 115: 1354–1363. doi: 10.1182/blood-2009-08-237370
|
| [55] |
Vassena L, Giuliani E, Matusali G, et al. (2013) The human immunodeficiency virus type 1 Vpr protein upregulates PVR via activation of the ATR-mediated DNA damage response pathway. J Gen Virol 94: 2664–2669. doi: 10.1099/vir.0.055541-0
|
| [56] |
Gasser S, Orsulic S, Brown EJ, et al. (2005) The DNA damage pathway regulates innate immune system ligands of the NKG2D receptor. Nature 436: 1186–1190. doi: 10.1038/nature03884
|
| [57] |
Soriani A, Zingoni A, Cerboni C, et al. (2009) ATM-ATR-dependent up-regulation of DNAM-1 and NKG2D ligands on multiple myeloma cells by therapeutic agents results in enhanced NK-cell susceptibility and is associated with a senescent phenotype. Blood 113: 3503–3511. doi: 10.1182/blood-2008-08-173914
|
| [58] |
Croxford JL, Tang ML, Pan MF, et al. (2013) ATM-dependent spontaneous regression of early Eμ-myc-induced murine B-cell leukemia depends on natural killer and T cells. Blood 121: 2512–2521. doi: 10.1182/blood-2012-08-449025
|
| [59] |
Fionda C, Abruzzese MP, Zingoni AS, et al. (2015) Nitric oxide donors increase PVR/CD155 DNAM-1 ligand expression in multiple myeloma cells: role of DNA damage response activation. BMC Cancer 15: 17. doi: 10.1186/s12885-015-1023-5
|
| [60] |
Champsaur M, Lanier LL (2010) Effect of NKG2D ligand expression on host immune responses. Immunol Rev 235: 267–285. doi: 10.1111/j.0105-2896.2010.00893.x
|
| [61] |
Kamran N, Takai Y, Miyoshi J, et al. (2013) Toll-like receptor ligands induce expression of the costimulatory molecule CD155 on antigen-presenting cells. PLoS One 8: e54406. doi: 10.1371/journal.pone.0054406
|
| [62] |
Pignoloni B, Fionda C, Dell'Oste V, et al. (2016) Distinct roles for human cytomegalovirus immediate early proteins IE1 and IE2 in the transcriptional regulation of MICA and PVR/CD155 Expression. J Immunol 197: 4066–4078. doi: 10.4049/jimmunol.1502527
|
| [63] |
Solecki DJ, Gromeier M, Mueller S, et al. (2002) Expression of the human poliovirus receptor/CD155 gene is activated by sonic hedgehog. Biol Chem 277: 25697–25702. doi: 10.1074/jbc.M201378200
|
| [64] |
Hirota T, Irie K, Okamoto R, et al. (2005) Transcriptional activation of the mouse Necl-5/Tage4/PVR/CD155 gene by fibroblast growth factor or oncogenic Ras through the Raf-MEK-ERK-AP-1 pathway. Oncogene 24: 2229–2235. doi: 10.1038/sj.onc.1208409
|
| [65] |
Jung H, Hsiung B, Pestal K, et al. (2012) RAE-1 ligands for the NKG2D receptor are regulated by E2F transcription factors, which control cell cycle entry. J Exp Med 209: 2409–2422. doi: 10.1084/jem.20120565
|
| [66] |
Soriani A, Iannitto ML, Ricci B, et al. (2014) Reactive oxygen species- and DNA damage response-dependent NK cell activating ligand upregulation occurs at transcriptional levels and requires the transcriptional factor E2F1. J Immunol 193: 950–960. doi: 10.4049/jimmunol.1400271
|
| [67] |
Stern-Ginossar N, Gur C, Biton M, et al. (2008) Human microRNAs regulate stress-induced immune responses mediated by the receptor NKG2D. Nat Immunol 9: 1065–1073. doi: 10.1038/ni.1642
|
| [68] |
Yadav D, Ngolab J, Lim RS, et al. (2009) Cutting edge: down-regulation of MHC class I-related chain A on tumor cells by IFN-gamma-induced microRNA. J Immunol 182: 39–43. doi: 10.4049/jimmunol.182.1.39
|
| [69] |
Nachmani D, Stern-Ginossar N, Sarid R, et al. (2009) Diverse herpesvirus microRNAs target the stress-induced immune ligand MICB to escape recognition by natural killer cells. Cell Host Microbe 5: 376–385. doi: 10.1016/j.chom.2009.03.003
|
| [70] |
Heinemann A, Zhao F, Pechlivanis S, et al. (2012) Tumor suppressive microRNAs miR-34a/c control cancer cell expression of ULBP2, a stress-induced ligand of the natural killer cell receptor NKG2D. Cancer Res 72: 460–471. doi: 10.1158/0008-5472.CAN-11-1977
|
| [71] |
Boutet P, Agüera-González S, Atkinson S, et al. (2009) Cutting edge: the metalloproteinase ADAM17/TNF-alpha-converting enzyme regulates proteolytic shedding of the MHC class I-related chain B protein. J Immunol 182: 49–53. doi: 10.4049/jimmunol.182.1.49
|
| [72] |
Waldhauer I, Steinle A (2006) Proteolytic release of soluble UL16-binding protein 2 from tumor cells. Cancer Res 66: 2520–2526. doi: 10.1158/0008-5472.CAN-05-2520
|
| [73] |
Zingoni A, Cecere F, Vulpis E, et al. (2015) Genotoxic stress induces senescence-associated ADAM10-dependent release of NKG2D MIC ligands in multiple myeloma cells. J Immunol 195: 736–748. doi: 10.4049/jimmunol.1402643
|
| [74] |
Ashiru O, Boutet P, Fernández-Messina L, et al. (2010) Natural killer cell cytotoxicity is suppressed by exposure to the human NKG2D ligand MICA*008 that is shed by tumor cells in exosomes. Cancer Res 70: 481–489. doi: 10.1158/0008-5472.CAN-09-1688
|
| [75] |
Fernández-Messina L, Ashiru O, Boutet P, et al. (2010) Differential mechanisms of shedding of the glycosylphosphatidylinositol (GPI)-anchored NKG2D ligands. J Biol Chem 285: 8543–8551. doi: 10.1074/jbc.M109.045906
|
| [76] |
Iguchi-Manaka A, Okumura G, Kojima H, et al. (2016) Increased soluble CD155 in the serum of cancer patients. PLoS One 11: e0152982. doi: 10.1371/journal.pone.0152982
|
| [77] |
Ashiru O, Bennett NJ, Boyle LH, et al. (2009) NKG2D ligand MICA is retained in the cis-Golgi apparatus by human cytomegalovirus protein UL142. J Virol 83: 12345–12354. doi: 10.1128/JVI.01175-09
|
| [78] |
Bennett NJ, Ashiru O, Morgan FJ, et al. (2010) Intracellular sequestration of the NKG2D ligand ULBP3 by human cytomegalovirus. J Immunol 185: 1093–10102. doi: 10.4049/jimmunol.1000789
|
| [79] |
Prod'Homme V, Sugrue DM, Stanton RJ, et al. (2010) Human cytomegalovirus UL141 promotes efficient downregulation of the natural killer cell activating ligand CD112. J Gen Virol 91: 2034–2039. doi: 10.1099/vir.0.021931-0
|
| [80] |
Schepis D, D'Amato M, Studahl M, et al. (2009) Herpes simplex virus infection downmodulates NKG2D ligand expression. Scand J Immunol 69: 429–436. doi: 10.1111/j.1365-3083.2009.02241.x
|
| [81] |
Gong J, Fang L, Liu R, et al. (2014) UPR decreases CD226 ligand CD155 expression and sensitivity to NK cell-mediated cytotoxicity in hepatoma cells. Eur J Immunol 44: 3758–3767. doi: 10.1002/eji.201444574
|
| [82] | Bologna S, Ferrari S (2013) It takes two to tango: ubiquitin and SUMO in the DNA damage response. Front Genet 4: 106. |
| [83] |
Wimmer P, Schreiner S (2015) Viral mimicry to usurp ubiquitin and SUMO host pathways. Viruses 7: 4854–4872. doi: 10.3390/v7092849
|
| [84] |
Ardorisio S, Fierabracci A, Muscari I, et al. (2017) SUMO proteins: guardians of immune system. J Autoimmun 84: 21–28. doi: 10.1016/j.jaut.2017.09.001
|
| [85] |
Thomas M, Boname JM, Field S, et al. (2008) Down-regulation of NKG2D and NKp80 ligands by Kaposi's sarcoma-associated herpesvirus K5 protects against NK cell cytotoxicity. Proc Natl Acad Sci USA 105: 1656–1661. doi: 10.1073/pnas.0707883105
|
| [86] |
Nice TJ, Coscoy L, Raulet DH (2009) Posttranslational regulation of the NKG2D ligand Mult1 in response to cell stress. J Exp Med 206: 287–298. doi: 10.1084/jem.20081335
|
| [87] |
Nice TJ, Deng W, Coscoy L, et al. (2010) Stress-regulated targeting of the NKG2D ligand Mult1 by a membrane-associated RING-CH family E3 ligase. J Immunol 185: 5369–7536. doi: 10.4049/jimmunol.1000247
|
| [88] |
Zitti B, Molfetta R, Fionda C, et al. (2017) Innate immune activating ligand SUMOylation affects tumor cell recognition by NK cells. Sci Rep 7: 10445. doi: 10.1038/s41598-017-10403-0
|
| [89] |
Komander D, Rape M (2012) The ubiquitin code. Annu Rev Biochem 81: 203–229. doi: 10.1146/annurev-biochem-060310-170328
|
| [90] |
Varshavsky A (2017) The ubiquitin system, autophagy, and regulated protein degradation. Annu Rev Biochem 86: 123–128. doi: 10.1146/annurev-biochem-061516-044859
|
| [91] |
Molfetta R, Gasparrini F, Santoni A, et al. (2010) Ubiquitination and endocytosis of the high affinity receptor for IgE. Mol Immunol 47: 2427–2434. doi: 10.1016/j.molimm.2010.06.003
|
| [92] |
Wilkinson KA, Henley JM (2010) Mechanisms, regulation and consequences of protein SUMOylation. Biochem J 428: 133–145. doi: 10.1042/BJ20100158
|
| [93] |
Flotho A, Melchior F (2013) Sumoylation: a regulatory protein modification in health and disease. Annu Rev Biochem 82: 357–385. doi: 10.1146/annurev-biochem-061909-093311
|
| [94] |
Fuertes MB, Girart MV, Molinero LL, et al. (2008) Intracellular retention of the NKG2D ligand MHC class I chain-related gene A in human melanomas confers immune privilege and prevents NK cell-mediated cytotoxicity. J Immunol 180: 4606–4614. doi: 10.4049/jimmunol.180.7.4606
|
| [95] |
Agüera-González S, Boutet P, Reyburn HT, et al. (2009) Brief residence at the plasma membrane of the MHC class I-related chain B is due to clathrin-mediated cholesterol-dependent endocytosis and shedding. J Immunol 182: 4800–4908. doi: 10.4049/jimmunol.0800713
|
| [96] |
Fernández-Messina L, Reyburn HT, Valés-Gómez M (2016) A short half-life of ULBP1 at the cell surface due to internalization and proteosomal degradation. Immunol Cell Biol 94: 479–485. doi: 10.1038/icb.2016.2
|
| [97] |
Castriconi R, Cantoni C, Della CM, et al. (2003) Transforming growth factor beta 1 inhibits expression of NKp30 and NKG2D receptors: consequences for the NK-mediated killing of dendritic cells. Proc Natl Acad Sci USA 100: 4120–4125. doi: 10.1073/pnas.0730640100
|
| [98] |
Lee JC, Lee KM, Kim DW, et al. (2004) Elevated TGF-beta1 secretion and down-modulation of NKG2D underlies impaired NK cytotoxicity in cancer patients. J Immunol 172: 7335–7340. doi: 10.4049/jimmunol.172.12.7335
|
| [99] |
Dasgupta S, Bhattacharya-Chatterjee M, Jr OB, et al. (2005) Inhibition of NK cell activity through TGF-beta 1 by down-regulation of NKG2D in a murine model of head and neck cancer. J Immunol 175: 5541–5550. doi: 10.4049/jimmunol.175.8.5541
|
| [100] |
Burgess SJ, Marusina AI, Pathmanathan I, et al. (2006) IL-21 down-regulates NKG2D/DAP10 expression on human NK and CD8+ T cells. J Immunol 176: 1490–1497. doi: 10.4049/jimmunol.176.3.1490
|
| [101] |
Zhang C, Zhang J, Niu J, et al. (2008) Interleukin-12 improves cytotoxicity of natural killer cells via upregulated expression of NKG2D. Hum Immunol 69: 490–500. doi: 10.1016/j.humimm.2008.06.004
|
| [102] |
Crane CA, Han SJ, Barry JJ, et al. (2010) TGF-beta downregulates the activating receptor NKG2D on NK cells and CD8+ T cells in glioma patients. Neuro Oncol 12: 7–13. doi: 10.1093/neuonc/nop009
|
| [103] |
Muntasell A, Magri G, Pende D, et al. (2010) Inhibition of NKG2D expression in NK cells by cytokines secreted in response to human cytomegalovirus infection. Blood 115: 5170–5179. doi: 10.1182/blood-2009-11-256479
|
| [104] |
Park YP, Choi SC, Kiesler P, et al. (2011) Complex regulation of human NKG2D-DAP10 cell surface expression: opposing roles of the γc cytokines and TGF-β1. Blood 118: 3019–3027. doi: 10.1182/blood-2011-04-346825
|
| [105] |
Groh V, Wu J, Yee C, et al. (2002) Tumor-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature 419: 734–738. doi: 10.1038/nature01112
|
| [106] |
Doubrovina ES, Doubrovin MM, Vider E, et al. (2003) Evasion from NK cell immunity by MHC class I chain-related molecules expressing colon adenocarcinoma. J Immunol 171: 6891–6899. doi: 10.4049/jimmunol.171.12.6891
|
| [107] |
Ogasawara K, Hamerman JA, Hsin H, et al. (2003) Impairment of NK cell function by NKG2D modulation in NOD mice. Immunity 18: 41–51. doi: 10.1016/S1074-7613(02)00505-8
|
| [108] |
Oppenheim DE, Roberts SJ, Clarke SL, et al. (2005) Sustained localized expression of ligand for the activating NKG2D receptor impairs natural cytotoxicity in vivo and reduces tumor immunosurveillance. Nat Immunol 6: 928–937. doi: 10.1038/ni1239
|
| [109] |
Wiemann K, Mittrücker HW, Feger U, et al. (2005) Systemic NKG2D down-regulation impairs NK and CD8 T cell responses in vivo. J Immunol 175: 720–729. doi: 10.4049/jimmunol.175.2.720
|
| [110] |
Cerboni C, Ardolino M, Santoni A, et al. (2009) Detuning CD8+ T lymphocytes by down-regulation of the activating receptor NKG2D: role of NKG2D ligands released by activated T cells. Blood 113: 2955–2964. doi: 10.1182/blood-2008-06-165944
|
| [111] |
Molfetta R, Quatrini L, Capuano C, et al. (2014) c-Cbl regulates MICA- but not ULBP2-induced NKG2D down-modulation in human NK cells. Eur J Immunol 44: 2761–2770. doi: 10.1002/eji.201444512
|
| [112] |
Molfetta R, Quatrini L, Zitti B, et al. (2016) Regulation of NKG2D expression and signaling by endocytosis. Trends Immunol 37: 790–802. doi: 10.1016/j.it.2016.08.015
|
| [113] |
Roda-Navarro P, Reyburn HT (2009) The traffic of the NKG2D/Dap10 receptor complex during natural killer (NK) cell activation. J Biol Chem 284: 16463–16472. doi: 10.1074/jbc.M808561200
|
| [114] |
Quatrini L, Molfetta R, Zitti B, et al. (2015) Ubiquitin-dependent endocytosis of NKG2D-DAP10 receptor complexes activates signaling and functions in human NK cells. Sci Signal 8: ra108. doi: 10.1126/scisignal.aab2724
|
| [115] |
Horng T, Bezbradica JS, Medzhitov R (2007) NKG2D signaling is coupled to the interleukin 15 receptor signaling pathway. Nat Immunol 8: 1345–1352. doi: 10.1038/ni1524
|
| [116] |
Paolini R, Serra A, Molfetta R, et al. (1999) Tyrosine kinase-dependent ubiquitination of CD16 zeta subunit in human NK cells following receptor engagement. Eur J Immunol 29: 3179–3187. doi: 10.1002/(SICI)1521-4141(199910)29:10<3179::AID-IMMU3179>3.0.CO;2-9
|
| [117] |
Paolini R, Molfetta R, Piccoli M, et al. (2001) Ubiquitination and degradation of Syk and ZAP-70 protein tyrosine kinases in human NK cells upon CD16 engagement. Proc Natl Acad Sci USA 98: 9611–9616. doi: 10.1073/pnas.161298098
|
| [118] |
Capuano C, Romanelli M, Pighi C, et al. (2015) Anti-CD20 therapy acts via FcγRIIIA to diminish responsiveness of human natural killer cells. Cancer Res 75: 4097–4108. doi: 10.1158/0008-5472.CAN-15-0781
|
| [119] |
Capuano C, Pighi C, Molfetta R, et al. (2017) Obinutuzumab-mediated high-affinity ligation of FcγRIIIA/CD16 primes NK cells for IFNγ production. Oncoimmunology 6: e1290037. doi: 10.1080/2162402X.2017.1290037
|
| [120] |
Wilson EB, El-Jawhari JJ, Neilson AL, et al. (2011) Human tumour immune evasion via TGF-β blocks NK cell activation but not survival allowing therapeutic restoration of anti-tumour activity. PLoS One 6: e22842. doi: 10.1371/journal.pone.0022842
|
| [121] |
Sanchez-Correa B, Gayoso I, Bergua JM, et al. (2012) Decreased expression of DNAM-1 on NK cells from acute myeloid leukemia patients. Immunol Cell Biol 90: 109–115. doi: 10.1038/icb.2011.15
|
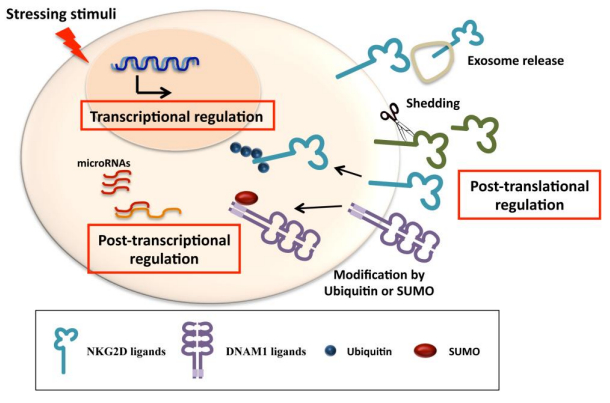
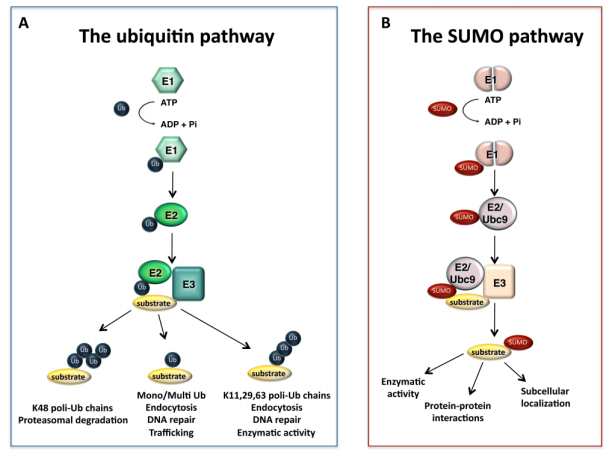
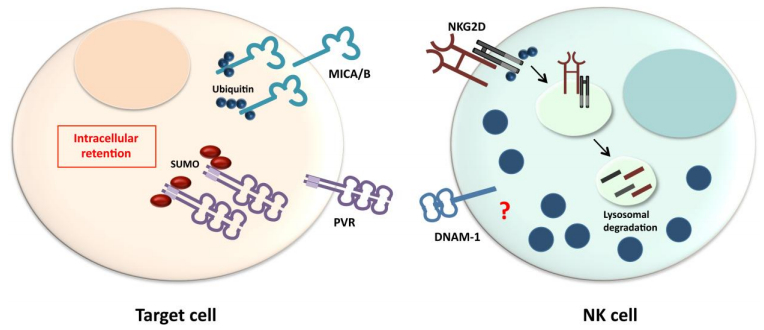
Figures(3)

Rosa Molfetta, Beatrice Zitti, Angela Santoni, Rossella Paolini. Ubiquitin and ubiquitin-like modifiers modulate NK cell-mediated recognition and killing of damaged cells[J]. AIMS Allergy and Immunology, 2017, 1(4): 164-180. doi: 10.3934/Allergy.2017.4.164







 DownLoad:
DownLoad: