Citation: María Santos-Corrada, Rafael Méndez-Tejeda. Public perception of climate change in a period of economic crisis in Puerto Rico[J]. AIMS Environmental Science, 2017, 4(1): 83-93. doi: 10.3934/environsci.2017.1.83

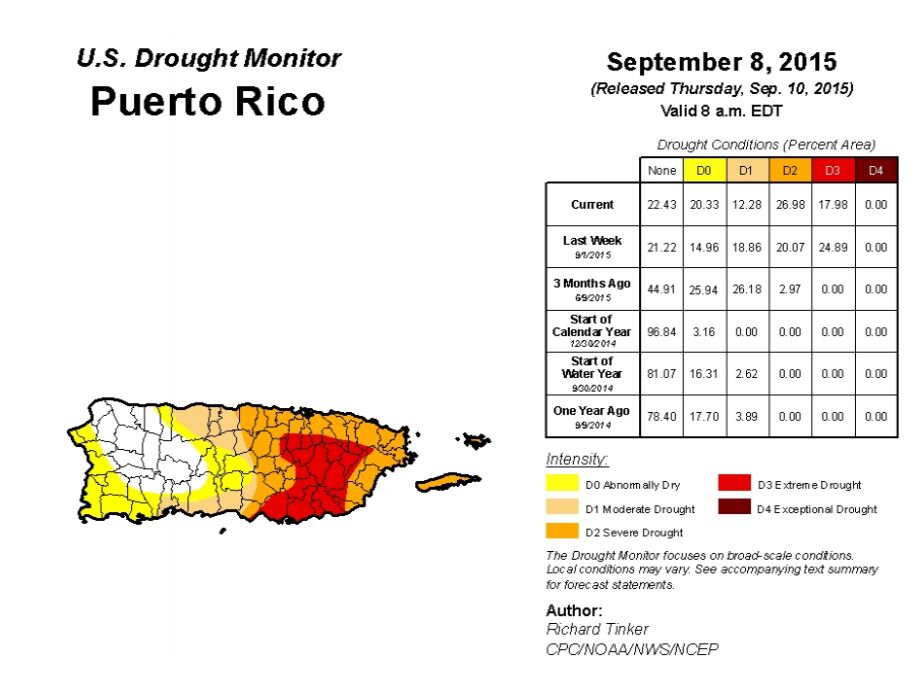
| [1] | The national drougt mitigation center, available from: http://droughtmonitor.unl.edu/Home/StateDroughtMonitor.aspx?PR |
| [2] |
Papoulis D, Kaika D, Bampatsou C, et al. (2015) Public Perception of Climate Change in a Period of Economic Crisis. Climate 3: 715-726. doi: 10.3390/cli3030715

|
| [3] | Symon Carolyn (2005) Artic Climate Impact Assessment. Cambridge University Press. 1042p. |
| [4] |
Pielke Sr RA (2005) Land Use and Climate Change. Science 310: 1625-1626. doi: 10.1126/science.1120529
|
| [5] | PRCCC. Puerto Rico's State of the Climate 2010-2013 Assessing Puerto Rico's Social-Ecological Vulnerabilities in a Changing Climate, 2013. Available from: http://pr-ccc.org/download/PR%20State%20of%20the%20Climate-FINAL_ENE2015.pdf |
| [6] |
Rafael MT, María SC, Sergio OM, et al. (2015) Environmental and Economic Impact of Forest Fires in Puerto Rico 2013-2014. Open J Forestry 5: 353. doi: 10.4236/ojf.2015.54030
|
| [7] | Atiles-Osoria JM (2014) Environmental Colonialism, Criminalization and Resistance: Puerto Rican Mobilizations for Environmental Justice in the 21st Century. RCCS Annual Review. A selection from the Portuguese journal Revista Crítica de Ciências Sociais. |
| [8] | Chisari O, Galiani S, Miller S (2013) Optimal Climate Change Adaptation and Mitigation in Environmentally Small Economies. IDB Working Paper No. 417. Inter-American Development Bank, Washington, DC. |
| [9] | Censo de Puerto Rico 2010, 2012. Available from: https://www.census.gov/prod/cen2010/cph-2-53sp.pdf. |
| [10] | Pagan-Trinidad I, Statistical Analyses of Spatial and Temporal Storm Rainfall Characteristics in Puerto Rico; Project No. 374103 Final Technical Report; United States Department of Interior: Mayaguez, PR, USA, 1984. |
| [11] |
Torres-Valcárcel A, Harbor J, González-Aviles C, et al. (2014) Impacts of Urban Development on Precipitation in the Tropical Maritime Climate of Puerto Rico. Climate 2: 47-77. doi: 10.3390/cli2020047
|
| [12] | Méndez-Lázaro PA, Nieves-Santiago A, Miranda-Bermúdez J (2014) Trends in Total Rainfall, Heavy Rain Events, and Number of Dry Days in San Juan, Puerto Rico, 1955-2009. Ecol Soc 2: 50. |
| [13] |
Rigau-Perez JG, Clark GG, Gubler DJ, et al. (1998) Dengue and Dengue Hemorrhagic Fever. Lancet 352: 971-977. doi: 10.1016/S0140-6736(97)12483-7
|
| [14] | Rigau-Perez JG, Ayala-Lopez A, Garcia-Rivera EJ, et al. (2002) The Reappearance of Dengue-3 and a Subsequent Dengue-4 and Dengue-1 Epidemic in Puerto Rico in 1998. ASTMH 67: 355-362. |
| [15] |
Méndez-Lázaro P, Muller-Karger FE, Otis D, et al. (2014). Assessing Climate Variability Effects on Dengue Incidence in San Juan, Puerto Rico. Int J Environ Res Public Health 11: 9409-9428. doi: 10.3390/ijerph110909409
|
| [16] | CDC. Zika in Puerto Rico a 'challenge and crisis'. Centers for Disease Control and Prevention (CDC). Available from: http://www.cdc.gov/. |
| [17] | Armstrong RA, Singh H, Torres J (2002) Benthic Survey of Insular Slope Coral Reefs using the SeaBed AUV. Backscatter 13: 22-25. |
| [18] | Ballantine DL, Appeldoorn RS, Yoshioka P, et al. (2008) Biology and ecology of Puerto Rican coral reefs. Coral Reefs of the USA, Springer Netherlands 2008: 375-406. |
| [19] | Hernández-Delgado EA (2000) Effects of Anthropogenic Stress Gradients in the Structure of Coral Reef Epibenthic and Fish Communities. Ph.D. Dissertation, Department of Biology, University of Puerto Rico, San Juan, 330 p. |
| [20] |
Irizarry-Soto E, Weil E (2009) Spatial and Temporal Variability in Juvenile Coral Densities, Survivorship and Recruitment in La Parguera, Southwestern Puerto Rico. Caribb J Sci 45: 269-281. doi: 10.18475/cjos.v45i2.a14
|
| [21] | Rafael MT, María SC, Sergio OM, et al. (2015) Environmental and Economic Impact of Forest Fires in Puerto Rico 2013-2014. Open J Forestry 5: 353. |
| [22] | PRCCC. Puerto Rico's State of the Climate 2010-2013: Assessing Puerto Rico's Social-Ecological Vulnerabilities in a Changing Climate. (2013). Available from: http://pr-ccc.org/download/PR%20State%20of%20the%20Climate-FINAL_ENE2015.pdf |
| [23] | Cabrera, Nahir, and Barreto, Maritza (2013) A Study of Beach Profile Changes at Selected Beaches on the North Coast of Puerto Rico (2009-2013). Geography Department, University of Puerto Rico, Río Piedras Campus Graduate Student, Graduate School of Planning, University Puerto Rico, Río Piedras Campus. Available from: http://cara.uprm.edu/?q=node/64/ |
| [24] | Van Beusekom AE, González G, Rivera MM (2014) Short-term Precipitation and Temperature Trends along an Elevation Gradient in Northeastern Puerto Rico. Earth Interactions 19: 1-33. |
| [25] | Carter M, Elsner J (1997) A Statistical Method for Forecasting Rainfall Over Puerto Rico. Wea Forecasting 12: 515–525. |
| [26] | García-Martinó AR, Warner GS, Scatena FN, et al. (1996). Rainfall, Runoff and Elevation Relationships in the Luquillo Mountains of Puerto Rico. Caribb J Sci 32: 413-424. |
| [27] | Larsen MC, Simon A (1993) A Rainfall Intensity-Duration Threshold for Landslides in a Humid-Tropical Environment, Puerto Rico. Geogr Ann 75A: 13-23. |
| [28] |
Johansson MA, Cummings DAT, Glass GE (2009) Multiyear Climate Variability and Dengue-El Niño Southern Oscillation, Weather, and Dengue Incidence in Puerto Rico, Mexico, and Thailand: A Longitudinal Data Analysis. PLoS Med 6: e1000168. doi: 10.1371/journal.pmed.1000168
|
| [29] |
Jury MR (2011) Caribbean Hurricanes: Interaction of Easterly and Westerly Waves. Theor Appl Climatol 106: 117-126. doi: 10.1007/s00704-011-0424-x
|
| [30] | Jury MR, Rios-Berrios R, Garcia E (2011) Caribbean Hurricanes: Changes of Intensity and Track Prediction. Theor Appl Climatol 107: 297-311. |
| [31] |
Donnelly JP, Woodruff JD (2007) Intense Hurricane Activity over the Past 5,000 Years. Nature 447: 465-468. doi: 10.1038/nature05834
|
| [32] |
Chen AA, Taylor MA (2002) Investigating the Link between Early Season Caribbean Rainfall and the El Niño +1 year. Int J Climatology 22: 87-106. doi: 10.1002/joc.711
|
| [33] | Chen AA, Roy A, McTavish J, et al. (1997) Using SST Anomalies to Predict Flood and Drought Conditions for the Caribbean. Center for Ocean-Land Atmosphere Studies. |
| [34] |
Elsner JB, Kossin JP, Jagger TH (2008) The increasing intensity of the strongest tropical cyclones. Nature 455: 92-95. doi: 10.1038/nature07234
|
| [35] | Mann ME, Emanuel KA (2006) Atlantic Hurricane Trends Linked to Climate Change. EOS, Transactions American Geophysical Union 87: 233-241. |
| [36] | NOAA. Available from: http://www.nhc.noaa.gov/data/publications.php#NOAA. |
| [37] | Impacto Económico del Huracán Georges en Puerto Rico. Available from: http://www.jp.gobierno.pr/Portal_JP/Portals/0/Publicaciones/PublicacionesHistoricasOnline/Impacto%20Econ%C3%B3mico%20del%20Hurac%C3%A1n%20Georges%20de%20PR%20-%20Abril%201999-2.pdf. |
| [38] | National hurricane center, Hurricanes in history. Avaible from: http://www.nhc.noaa.gov/outreach/history/ |
| [39] | Bueno R, Herzfeld C, Stanton AE, et al. (2008) The Caribbean and climate change. Available from: https://www.sei-international.org/mediamanager/documents/Publications/Climate/inaction-caribbean-es-eng.pdf. |
| [40] | Gros D, Puerto Rico and Greece: A Tale of Two Defaults in a Monetary Union. Centre for European Policy Studies, 2015. Available from: https://www.ceps.eu/system/files/HLB5_DG_PuertoRico_0.pdfGren. |
| [41] | Greene D, Puerto Rico's Battered Economy: The Greece of the Caribbean? 2013. Available from: http://www.npr.org/2013/02/06/171071377/puerto-ricos-battered-economy-the-greece-of-the-caribbean |
| [42] | United Nations Millennium Development Goals Indicators. Available from: http://datos.bancomundial.org/indicador/NY.GDP.PCAP.CD. |
| [43] | EPA. Environmental Protection Agency and the Municipality. Available from: https://www.epa.gov/newsreleases/municipality. |
Figures(1) / Tables(1)

María Santos-Corrada, Rafael Méndez-Tejeda. Public perception of climate change in a period of economic crisis in Puerto Rico[J]. AIMS Environmental Science, 2017, 4(1): 83-93. doi: 10.3934/environsci.2017.1.83







 DownLoad:
DownLoad: