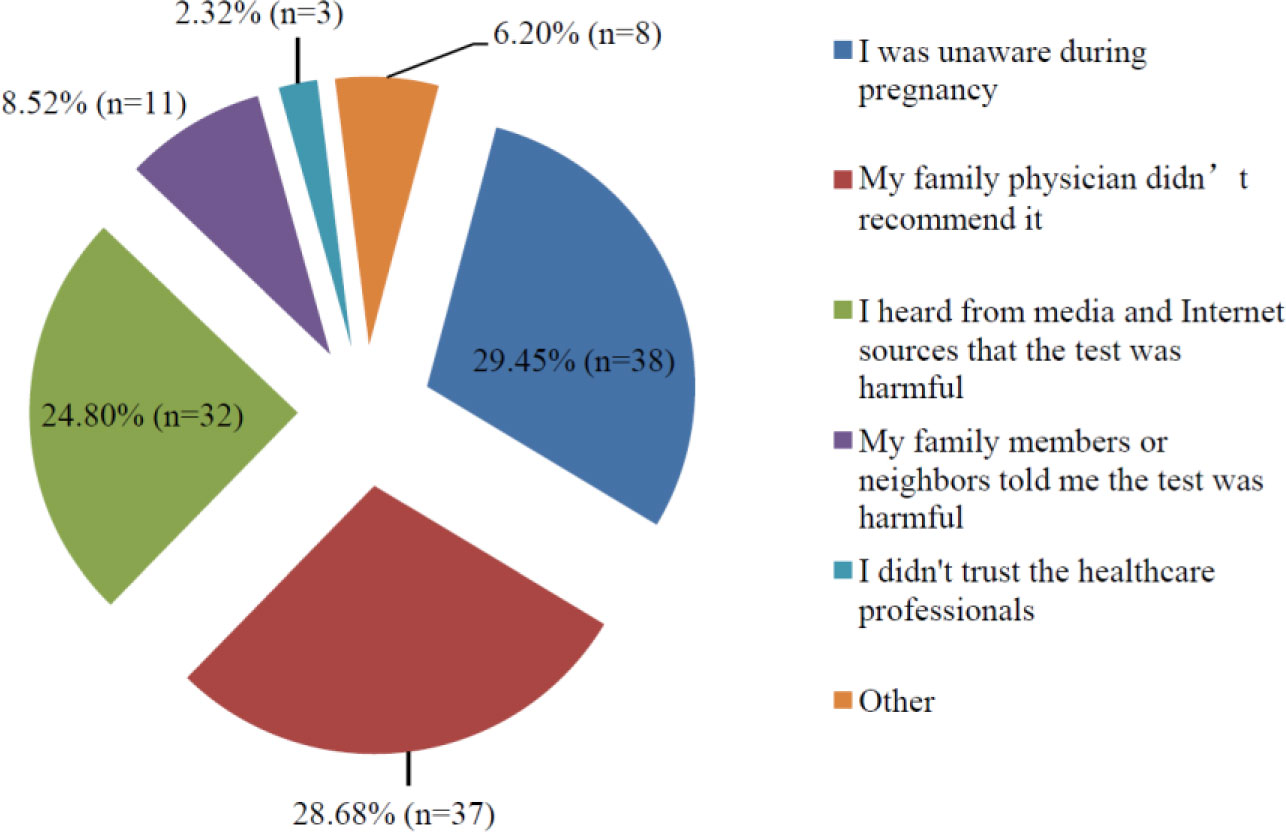
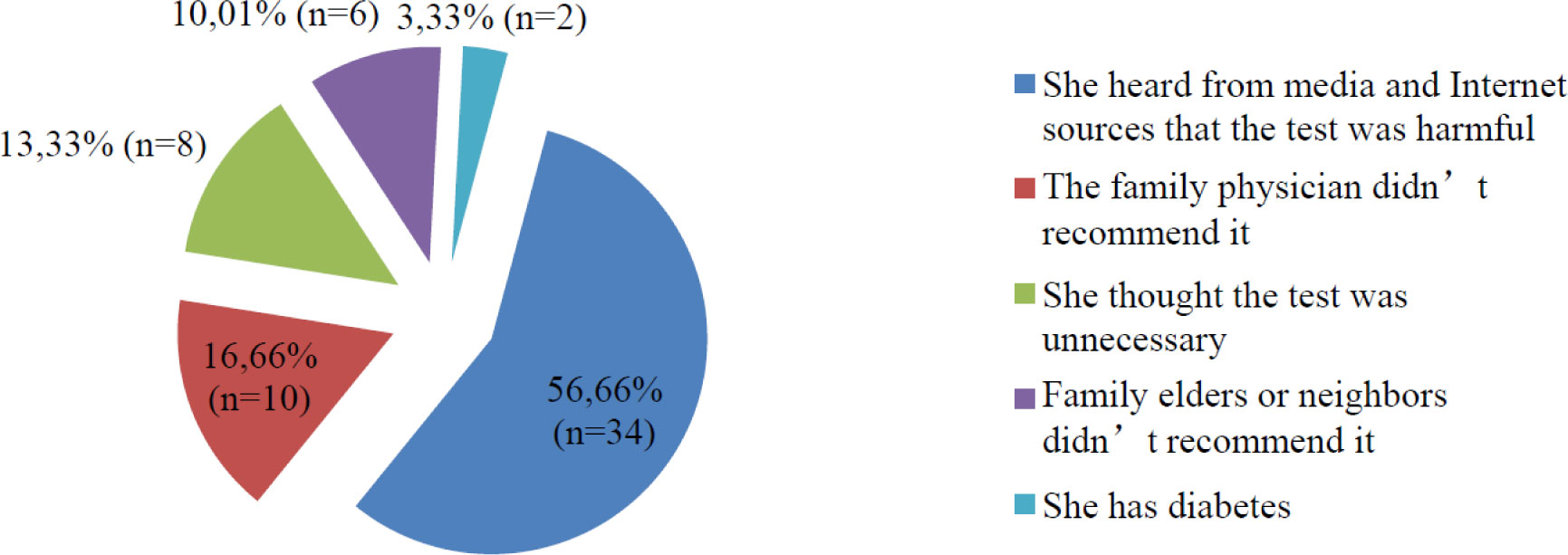
Discussions of the use of the oral glucose tolerance test (OGTT) took off when a Turkish scientist claimed in the media that “the OGTT is poisoning babies” in 2014. The aim of present study investigates the effects of controversies in the media and on the Internet on the attitudes and behaviors of women in regards to the OGTT. The research was designed as a descriptive, cross-sectional study. The universe of the study was women aged 18–45 years in Turkey. Included in the study were 358 women of childbearing age who attended family health center outpatient clinics in January 2019. A questionnaire was administered during face-to-face interviews to those who provided consent for participation in the study. The data was analyzed using SPSS 22.0 software. Chi-square test was used to compare the between-group qualitative data. The results were evaluated based on an alpha value of 0.05. Results: Of the participating women, 18.99% (n = 88) were unaware of the OGTT. Of participant, 41.89 (n = 151) delivered “Iwill take OGTT in the future”. 27.09% (n = 97) delivered “I will not” and 11.73% (n = 42) were hesitant. Of the participants, 67.32% (n = 241) reported having been pregnant in the past. Of the participant, 62.24% (n = 150) delivered that they had OGTT in the past. The reasons given for not undergoing the OGTT in past pregnancies were 29.45% (n = 38) unaware during pregnancy, 28.68% (n = 37) delivered “my family physician did not recommend it”. But the ones who delivered that in the future will not take OGTT, their reasons were 56.66% (n = 34) delivered “heard from media and internet sources” that the test was harmful. The results of the study indicate that public trust of OGTT and taking OGTT rates are declining in Turkey. While women can obtain beneficial information from media and Internet sources, misinformation can easily shake their confidence in any scientific data.
Citation: Gökmen Özceylan, Dilek Toprak. Effects of controversial statements on social media regarding the oral glucose tolerance testing on pregnant women in Turkey[J]. AIMS Public Health, 2020, 7(1): 20-28. doi: 10.3934/publichealth.2020003
Discussions of the use of the oral glucose tolerance test (OGTT) took off when a Turkish scientist claimed in the media that “the OGTT is poisoning babies” in 2014. The aim of present study investigates the effects of controversies in the media and on the Internet on the attitudes and behaviors of women in regards to the OGTT. The research was designed as a descriptive, cross-sectional study. The universe of the study was women aged 18–45 years in Turkey. Included in the study were 358 women of childbearing age who attended family health center outpatient clinics in January 2019. A questionnaire was administered during face-to-face interviews to those who provided consent for participation in the study. The data was analyzed using SPSS 22.0 software. Chi-square test was used to compare the between-group qualitative data. The results were evaluated based on an alpha value of 0.05. Results: Of the participating women, 18.99% (n = 88) were unaware of the OGTT. Of participant, 41.89 (n = 151) delivered “Iwill take OGTT in the future”. 27.09% (n = 97) delivered “I will not” and 11.73% (n = 42) were hesitant. Of the participants, 67.32% (n = 241) reported having been pregnant in the past. Of the participant, 62.24% (n = 150) delivered that they had OGTT in the past. The reasons given for not undergoing the OGTT in past pregnancies were 29.45% (n = 38) unaware during pregnancy, 28.68% (n = 37) delivered “my family physician did not recommend it”. But the ones who delivered that in the future will not take OGTT, their reasons were 56.66% (n = 34) delivered “heard from media and internet sources” that the test was harmful. The results of the study indicate that public trust of OGTT and taking OGTT rates are declining in Turkey. While women can obtain beneficial information from media and Internet sources, misinformation can easily shake their confidence in any scientific data.

| [1] | (2018) Turkey Association of Endocrine and Metabolism, diabetes mellitus and its complications diagnosis, treatment and monitoring guide.171. |
| [2] | Burtis CA, Ashwood ER, Bruns DE (2006) Tietz textbook of clinical chemistry and molecular diagnostics St Louis: Saunders, Elsevier, 498-499. |
| [3] |
Weinert LS (2010) International Association of Diabetes and Pregnancy Study Groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy: comment to the International Association of Diabetes and Pregnancy Study Groups Consensus Panel. Diabetes Care 33: e97-e97. doi: 10.2337/dc10-0544

|
| [4] | https://www.haberturk.com/saglik/haber/1012921-anne-karnindaki-bebegi-zehirliyorsunuz. |
| [5] | https://www.medimagazin.com.tr/hekim/genel/tr-tjod-baskanindan-karataya-hanimefendinin-herhangi-bir-calismasi-yok-2-12-67455.html. |
| [6] | https://doktorlarsitesi.net/2015/04/10/saglik-bakanligi-gebelere-seker-yukleme-testi-ile-ilgili-tartismalara-noktayi-koydu/. |
| [7] |
Griffin ME, Coffey M, Johnson H, et al. (2000) Universal vs. risk factor-based screening for gestational diabetes mellitus: detection rates, gestation at diagnosis and outcome. Diabetic Med 17: 26-32. doi: 10.1046/j.1464-5491.2000.00214.x
|
| [8] | Nakanishi S, Yoneda M, Maeda S (2013) Impact of glucose excursion and mean glucose concentration in oral glucose-tolerance test on oxidative stress among Japanese Americans. Diabetes, Metab Syndr Obes: Targets Ther 6: 427-433. |
| [9] |
Radovanovic PB, Kocijancic M (2015) How well are pregnant women in Croatia informed about the oral glucose tolerance test? Biochem Med 25: 230-236. doi: 10.11613/BM.2015.023
|
| [10] |
Eades CE, Cameron DM, Evans JMM (2017) Prevalence of gestational diabetes mellitus in Europe: A meta-analysis. Diabetes Res Clin Pract 129: 173-181. doi: 10.1016/j.diabres.2017.03.030
|
| [11] |
Gillani AH, Amirul İslam FM, Hayat K, et al. (2018) Knowledge, attitudes and practices regarding diabetes in the general population: A cross-sectional study from Pakistan. Int J Environ Res Public Health 15: 1906. doi: 10.3390/ijerph15091906
|
| [12] | Thani A, Farghaly AH, Akram H, et al. (2018) Public awareness and perceptions about diabetes in the State of Qatar. Cureus 10: e2671. |
| [13] |
Man B, Turyk ME, Kominiarek MA, et al. (2016) Diabetes screening in US women with a history of gestational diabetes, national health and nutrition examination survey, 2007–2012. Prev Chronic Dis 13: 160106. doi: 10.5888/pcd13.160106
|
| [14] |
Smirnakis KV, Chasan-Taber L, Wolf M, et al. (2005) Postpartum diabetes screening in women with a history of gestational diabetes. Obstet Gynecol 106: 1297-1303. doi: 10.1097/01.AOG.0000189081.46925.90
|
| [15] | Karasu Y (2018) What happened to the glucose loading test? The impact of media on public health. Med J Ankara Tr Res Hosp 51: 57-58. |
| [16] |
Rueangdetnarong H, Sekararithi R, Jaiwongkam T, et al. (2018) Comparisons of the oxidative stress biomarkers levels in GDM and non-GDM among Thai population: cohort study. Endocr Connect 7: 681-687. doi: 10.1530/EC-18-0093
|
| [17] |
Kassahun CH, Mekonen AG (2017) Knowledge, attitude, practices and their associated factors towards diabetes mellitus among non diabetes community members of Bale Zone administrative towns, South East Ethiopia. A cross-sectional study. Plos One 12: e0170040. doi: 10.1371/journal.pone.0170040
|
| [18] |
Karamanos B, Thanopoulou A, Anastasiou E, et al. (2014) Relation of the Mediterranean diet with the incidence of gestational diabetes. Eur J Clin Nutr 68: 8-13. doi: 10.1038/ejcn.2013.177
|
Figures(2) / Tables(5)

Gökmen Özceylan, Dilek Toprak. Effects of controversial statements on social media regarding the oral glucose tolerance testing on pregnant women in Turkey[J]. AIMS Public Health, 2020, 7(1): 20-28. doi: 10.3934/publichealth.2020003







 DownLoad:
DownLoad: