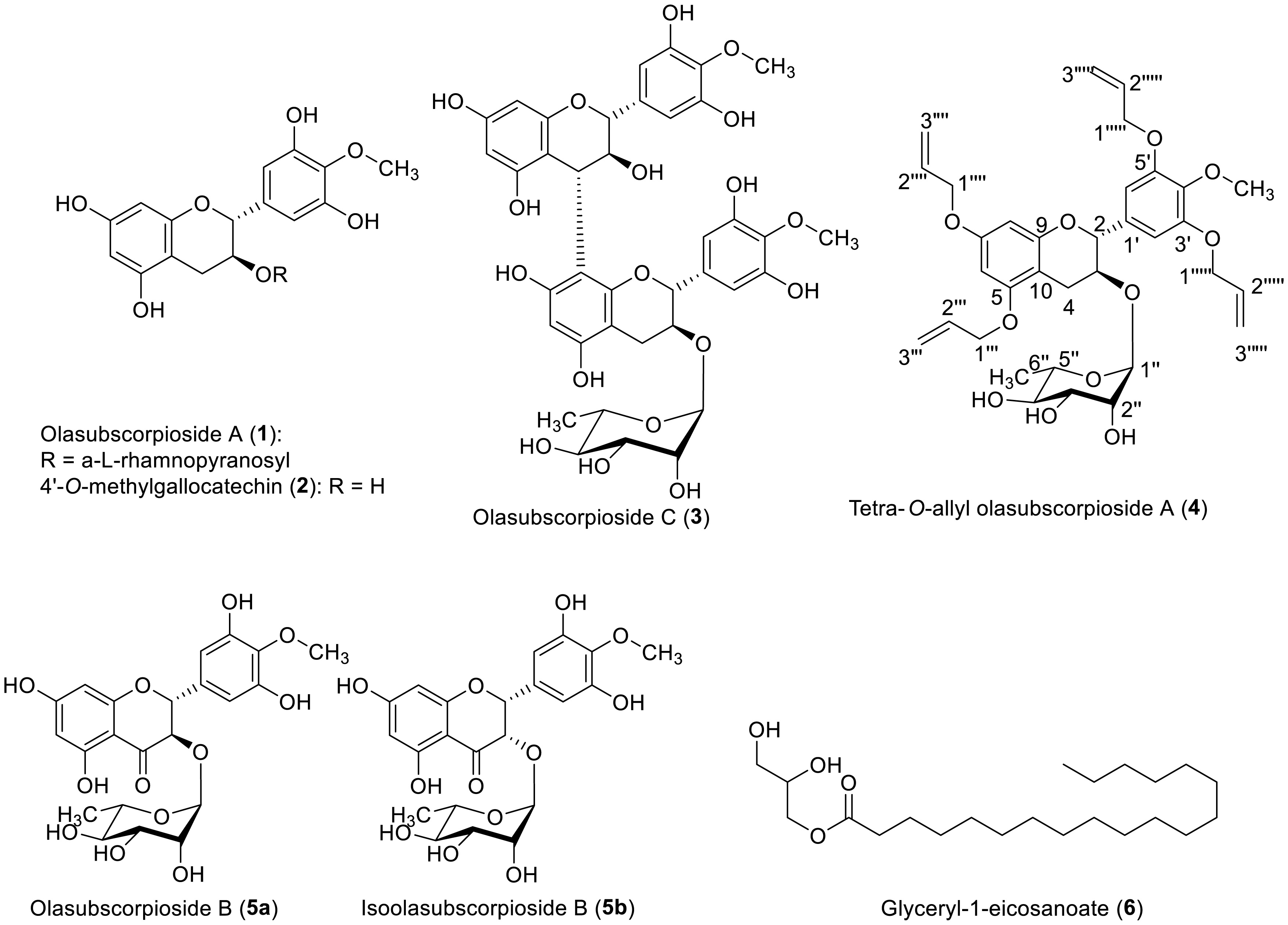
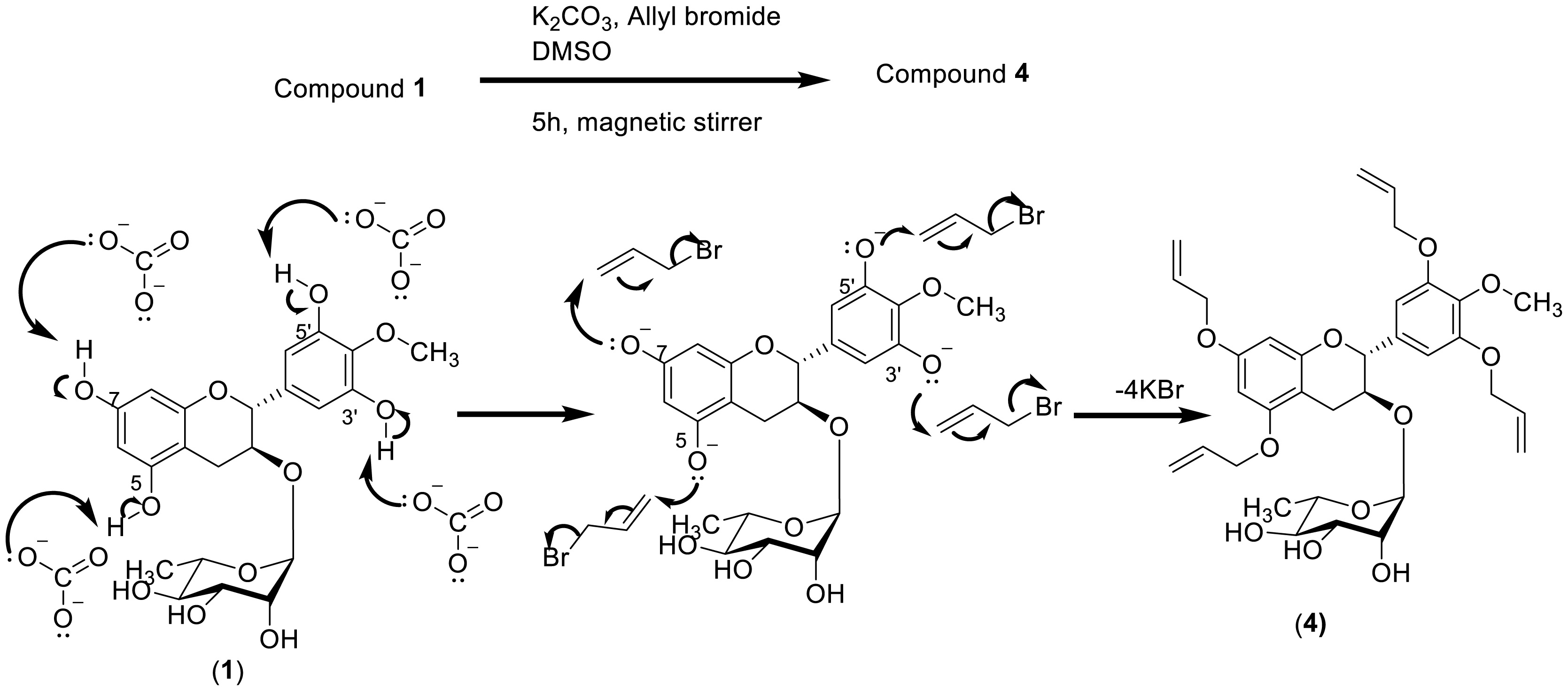
The recurrence of bacterial infections is commonly associated with the multiple interactions between humans and their environment. These interactions have progressively contributed to the development of antimicrobial resistance among numerous clinically important pathogenic bacteria. The present study targeted the antibacterial activity of natural products derived from the stem bark of O. subscorpioidea against clinical isolates of bacteria. This included the Gram-positive bacterium Staphylococcus aureus and three Gram-negative bacteria, Pseudomonas aeruginosa, Providencia stuartii, and Escherichia coli. The plant material was extracted by maceration, and chromatography was performed to isolate the chemical constituents from plant extracts as well as from the reaction medium. The resulting compounds were characterized using spectroscopic and spectrometric techniques. The antibacterial activity was assessed by the microdilution method. Five compounds were isolated, including olasubscorpioside A (1), 4′-O-methylgallocatechin (2), olasubscorpioside C (3), olasubscorpioside B (5a), isoolasubcorpioside B (5b), and glyceryl-1-eicosanoate (6). The semisynthetic derivatives 4′-O-methylgallocatechin (2) (also isolated from the plant extract) and tetra-O-allyl olasubscorpioside A (4) were prepared and characterized. Interestingly, olasubscorpioside A (1) and 4′-O-methylgallocatechin (2) displayed a significant antibacterial effect (MIC = 8 µg/mL) against the pathogenic isolate Staphylococcus aureus resistant to methicillin, ofloxacin, kanamycin, tetracycline, and erythromycin. Compound 2 had a better activity relative to 1 against Pseudomonas aeruginosa (from MIC > 256 to 128 µg/mL) and Escherichia coli (from MIC = 128 to 64 µg/mL). Gallocatechins from the stem bark of Olax subscorpioidea are promising bioactive molecules to fight against multiresistant bacteria.
Citation: Bienvenu Tsakem, Billy Toussie Tchegnitegni, Valaire Yemene Matieta, Xavier Siwe Noundou, Beaudelaire Kemvoufo Ponou, Syed Ghulam Musharraf, Léon Azefack Tapondjou, Victor Kuete, Rémy Bertrand Teponno. Gallocatechin analogues from Olax subscorpioidea Oliv. against multiresistant clinical bacterial isolates[J]. AIMS Molecular Science, 2025, 12(2): 173-186. doi: 10.3934/molsci.2025011
The recurrence of bacterial infections is commonly associated with the multiple interactions between humans and their environment. These interactions have progressively contributed to the development of antimicrobial resistance among numerous clinically important pathogenic bacteria. The present study targeted the antibacterial activity of natural products derived from the stem bark of O. subscorpioidea against clinical isolates of bacteria. This included the Gram-positive bacterium Staphylococcus aureus and three Gram-negative bacteria, Pseudomonas aeruginosa, Providencia stuartii, and Escherichia coli. The plant material was extracted by maceration, and chromatography was performed to isolate the chemical constituents from plant extracts as well as from the reaction medium. The resulting compounds were characterized using spectroscopic and spectrometric techniques. The antibacterial activity was assessed by the microdilution method. Five compounds were isolated, including olasubscorpioside A (1), 4′-O-methylgallocatechin (2), olasubscorpioside C (3), olasubscorpioside B (5a), isoolasubcorpioside B (5b), and glyceryl-1-eicosanoate (6). The semisynthetic derivatives 4′-O-methylgallocatechin (2) (also isolated from the plant extract) and tetra-O-allyl olasubscorpioside A (4) were prepared and characterized. Interestingly, olasubscorpioside A (1) and 4′-O-methylgallocatechin (2) displayed a significant antibacterial effect (MIC = 8 µg/mL) against the pathogenic isolate Staphylococcus aureus resistant to methicillin, ofloxacin, kanamycin, tetracycline, and erythromycin. Compound 2 had a better activity relative to 1 against Pseudomonas aeruginosa (from MIC > 256 to 128 µg/mL) and Escherichia coli (from MIC = 128 to 64 µg/mL). Gallocatechins from the stem bark of Olax subscorpioidea are promising bioactive molecules to fight against multiresistant bacteria.

| [1] |
Mancuso G, Midiri A, Gerace E, et al. (2021) Bacterial antibiotic resistance: The most critical pathogens. Pathogens 10: 1310. https://doi.org/10.3390/pathogens10101310 
|
| [2] |
Dance A (2024) Five ways science is tackling the antibiotic resistance crisis. Nature 632: 494-496. https://doi.org/10.1038/d41586-024-02601-4
|
| [3] |
Seukep AJ, Mbuntcha HG, Zeuko'o EM, et al. (2023) Established antibacterial drugs from plants. Advances in botanical research . Elsevier 81-149. https://doi.org/10.1016/bs.abr.2022.08.005
|
| [4] |
Keita K, Darkoh C, Okafor F (2022) Secondary plant metabolites as potent drug candidates against antimicrobial-resistant pathogens. SN Appl Sci 4: 209. https://doi.org/10.1007/s42452-022-05084-y
|
| [5] |
Heeb S, Fletcher MP, Chhabra SR, et al. (2011) Quinolones: From antibiotics to autoinducers. FEMS Microbiol Rev 35: 247-274. https://doi.org/10.1111/j.1574-6976.2010.00247.x
|
| [6] |
Tsakem B, Li G, Teponno RB (2024) Structures, biosynthesis and biological activities of benastatins, anthrabenzoxocinones and fredericamycins. Bioorg Chem 150: 107572. https://doi.org/10.1016/j.bioorg.2024.107572
|
| [7] |
Jain H, Chella N (2020) Solubility enhancement techniques for natural product delivery. Sustainable agriculture reviews . Cham: Springer 33-66. https://doi.org/10.1007/978-3-030-41838-0_2
|
| [8] | Tsakem B, Ponou BK, Toussie BT, et al. (2021) The Genus Olax. Natural products chemistry of botanical medicines from cameroonian plants . CRC Press 1-21. https://doi.org/10.1201/9780429506734-1 |
| [9] |
Tsakem B, Eckhardt P, Tchuenguem RT, et al. (2022) Muellerilactone and other bioactive constituents of Phyllanthus muellerianus (kuntze) exell. Biochem Syst Ecol 101: 104397. https://doi.org/10.1016/j.bse.2022.104397
|
| [10] |
Swana L, Tsakem B, Tembu JV, et al. (2023) The genus dacryodes Vahl.: Ethnobotany, Phytochemistry and Biological Activities. Pharmaceuticals 16: 775. https://doi.org/10.3390/ph16050775
|
| [11] |
Tsafack BT, Bomgning CLK, Kühlborn J, et al. (2018) Protective effects of extracts, isolated compounds from Desmodium uncinatum and semi-synthetic isovitexin derivatives against lipid peroxidation of hepatocyte's membranes. Adv Biol Chem 8: 101-120.
|
| [12] |
Tsakem B, Tchuenguem RT, Siwe-Noundou X, et al. (2022) New bioactive flavonoid glycosides with antioxidant activity from the stem bark of Olax subscorpioidea Oliv. Nat Prod Res 37: 1641-1650. https://doi.org/10.1080/14786419.2022.2106566
|
| [13] |
Garcia J, Massoma T, Morin C, et al. (1993) 4′-O-methylgallocatechin from Panda oleosa. Phytochemistry 32: 1626-1628. https://doi.org/10.1016/0031-9422(93)85196-X
|
| [14] |
Tsakem B, Toussie BT, Siwe-Noundou X, et al. (2023) Structure elucidation of olasubscorpioside C, a new rotameric biflavonoid glycoside from the stem barks of Olax subscorpioidea (Oliv). Magn Reson Chem 61: 497-503. https://doi.org/10.1002/mrc.5375
|
| [15] |
Darwish FMM, Reinecke MG (2003) Ecdysteroids and other constituents from Sida spinosa L. Phytochemistry 62: 1179-1184. https://doi.org/10.1016/S0031-9422(03)00021-9
|
| [16] |
Paudel A, Hamamoto H, Kobayashi Y, et al. (2012) Identification of novel deoxyribofuranosyl indole antimicrobial agents. J Antibiot 65: 53-57. https://doi.org/10.1038/ja.2011.110
|
| [17] | Dzoyem JP, Hamamoto H, Ngameni B, et al. (2013) Antimicrobial action mechanism of flavonoids from Dorstenia species. Drug Discov Ther 7: 66-72. |
| [18] |
Kuete V, Ngameni B, Tangmouo JG, et al. (2010) Efflux Pumps are involved in the defense of Gram-negative bacteria against the natural products isobavachalcone and diospyrone. Antimicrob Agents Chemother 54: 1749-1752. https://doi.org/10.1128/aac.01533-09
|
| [19] |
Lorenzi V, Muselli A, Bernardini AF, et al. (2009) Geraniol restores antibiotic activities against multidrug-resistant isolates from Gram-negative species. Antimicrob Agents Chemother 53: 2209-2211. https://doi.org/10.1128/aac.00919-08
|
| [20] |
Chevalier J, Pagès JM, Eyraud A, et al. (2000) Membrane permeability modifications are involved in antibiotic resistance in Klebsiella pneumoniae. Biochem Biophys Res Commun 274: 496-499. https://doi.org/10.1006/bbrc.2000.3159
|
| [21] |
Mativandlela SPN, Lall N, Meyer JJM (2006) Antibacterial, antifungal and antitubercular activity of (the roots of) Pelargonium reniforme (CURT) and Pelargonium sidoides (DC) (Geraniaceae) root extracts. S Afr J Bot 72: 232-237. https://doi.org/10.1016/j.sajb.2005.08.002
|
| [22] |
Garcia KYM, Quimque MTJ, Lambert C, et al. (2022) Antiproliferative and cytotoxic cytochalasins from Sparticola triseptata inhibit Actin polymerization and aggregation. J Fungi 8: 560. https://doi.org/10.3390/jof8060560
|
| [23] |
Kuete V (2010) Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Med 76: 1479-1491. https://doi.org/10.1055/s-0030-1250027
|
| [24] |
Botta B, Vitali A, Menendez P, et al. (2005) Prenylated flavonoids: Pharmacology and biotechnology. Curr Med Chem 12: 713-739. https://doi.org/10.2174/0929867053202241
|
| [25] |
Alhassan AM, Abdullahi MI, Uba A, et al. (2014) Prenylation of aromatic secondary metabolites: A new frontier for development of novel drugs. Trop J Pharm Res 13: 307. https://doi.org/10.4314/tjpr.v13i2.22
|
| [26] |
Fotso GW, Ngameni B, Storr TE, et al. (2020) Synthesis of novel stilbene–coumarin derivatives and antifungal screening of monotes kerstingii-specialized metabolites against Fusarium oxysporum. Antibiotics 9: 537. https://doi.org/10.3390/antibiotics9090537
|
| [27] | Patil SM, Patel P (2021) Bactericidal and bacteriostatic antibiotics. Infections and sepsis development . IntechOpen. https://doi.org/10.5772/intechopen.99546 |
| [28] |
Guefack MF, Messina NDM, Mbaveng AT, et al. (2022) Antibacterial and antibiotic-potentiation activities of the hydro-ethanolic extract and protoberberine alkaloids from the stem bark of against multidrug-resistant bacteria expressing active efflux pumps. J Ethnopharmacol 296: 115518. https://doi.org/10.1016/j.jep.2022.115518
|
| [29] |
Otsuka N, Liu MH, Shiota S, et al. (2008) Anti-methicillin resistant Staphylococcus aureus (MRSA) compounds isolated from Laurus nobilis. Biol Pharm Bull 31: 1794-1797.
|
| [30] |
Shamsudin NF, Ahmed QU, Mahmood S, et al. (2022) Antibacterial effects of flavonoids and their structure-activity relationship study: A comparative interpretation. Molecules 27: 1149. https://doi.org/10.3390/molecules27041149
|
| [31] |
Liu S, Zhang Q, Li H, et al. (2022) Comparative assessment of the antibacterial efficacies and mechanisms of different tea extracts. Foods 11: 620. https://doi.org/10.3390/foods11040620
|
| [32] |
Nguyen TLA, Bhattacharya D (2022) Antimicrobial activity of quercetin: An approach to its mechanistic principle. Molecules 27: 2494. https://doi.org/10.3390/molecules27082494
|
 molsci-12-02-011-s001.pdf molsci-12-02-011-s001.pdf |
 |
Figures(2) / Tables(2)

Bienvenu Tsakem, Billy Toussie Tchegnitegni, Valaire Yemene Matieta, Xavier Siwe Noundou, Beaudelaire Kemvoufo Ponou, Syed Ghulam Musharraf, Léon Azefack Tapondjou, Victor Kuete, Rémy Bertrand Teponno. Gallocatechin analogues from Olax subscorpioidea Oliv. against multiresistant clinical bacterial isolates[J]. AIMS Molecular Science, 2025, 12(2): 173-186. doi: 10.3934/molsci.2025011







 DownLoad:
DownLoad: