The cancer cells could be celled biomass without normal cellular regulation. They bypass most of the signaling pathways leading to programmed cell division. On the other hand, the embryos are highly regulated, giving rise to the whole organism based on the planned regulation. Understanding the bridge concepts between them might be an interventional art for discovering valuable cancer drugs. The present review highlighted the most similarities between them and recent literary works.
Citation: Kishore Kumar Meenakshi Sundaram, Giridharan Bupesh, Konda Mani Saravanan. Instrumentals behind embryo and cancer: a platform for prospective future in cancer research[J]. AIMS Molecular Science, 2022, 9(1): 25-45. doi: 10.3934/molsci.2022002
The cancer cells could be celled biomass without normal cellular regulation. They bypass most of the signaling pathways leading to programmed cell division. On the other hand, the embryos are highly regulated, giving rise to the whole organism based on the planned regulation. Understanding the bridge concepts between them might be an interventional art for discovering valuable cancer drugs. The present review highlighted the most similarities between them and recent literary works.

| [1] |
Blagosklonny MV (2005) Teratogens as anti-cancer drugs. Cell Cycle 4: 1518-1521. https://doi.org/10.4161/cc.4.11.2208 
|
| [2] |
Ramazani M, Jaktaji RP, Shirazi FH, et al. (2019) Analysis of apoptosis related genes in nurses exposed to anti-neoplastic drugs. BMC Pharmacol Toxicol 20. https://doi.org/10.1186/s40360-019-0372-0
|
| [3] |
Finnell RH (1999) Teratology: general considerations and principles. J Allergy Clin Immunol 103: S337-S342. https://doi.org/10.1016/S0091-6749(99)70259-9
|
| [4] |
Vargesson N (2015) Thalidomide-induced teratogenesis: History and mechanisms. Birth Defects Res Part C Embryo Today Rev 105: 140-156. https://doi.org/10.1002/bdrc.21096
|
| [5] |
Coyle I, Wayner MJ, Singer G (1976) Behavioral teratogenesis: A critical evaluation. Pharmacol Biochem Behav 4: 191-200. https://doi.org/10.1016/0091-3057(76)90014-9
|
| [6] |
Flaxman SM, Sherman PW (2000) Morning sickness: a mechanism for protecting mother and embryo. Q Rev Biol 75: 113-148. https://doi.org/10.1086/393377
|
| [7] |
Mei N, Guo X, Ren Z, et al. (2017) Review of Ginkgo biloba-induced toxicity, from experimental studies to human case reports. J Environ Sci Heal Part C 35: 1-28. https://doi.org/10.1080/10590501.2016.1278298
|
| [8] |
Wu C-P, Ohnuma S, V Ambudkar S (2011) Discovering natural product modulators to overcome multidrug resistance in cancer chemotherapy. Curr Pharm Biotechnol 12: 609-620. https://doi.org/10.2174/138920111795163887
|
| [9] |
Sung H, Ferlay J, Siegel RL, et al. (2021) Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 71: 209-249. https://doi.org/10.3322/caac.21660
|
| [10] | Roy M, Datta A (2019) Cancer: Types and Hallmarks. Cancer Genetics and Therapeutics . Springer 1-26. https://doi.org/10.1007/978-981-13-9471-3_1 |
| [11] |
Manzo G (2019) Similarities Between Embryo Development and Cancer Process Suggest New Strategies for Research and Therapy of Tumors: A New Point of View. Front cell Dev Biol 7: 20. https://doi.org/10.3389/fcell.2019.00020
|
| [12] |
Shabo I, Svanvik J, Lindström A, et al. (2020) Roles of cell fusion, hybridization and polyploid cell formation in cancer metastasis. World J Clin Oncol 11: 121. https://doi.org/10.5306/wjco.v11.i3.121
|
| [13] |
Ledbetter DH (2009) Chaos in the embryo. Nat Med 15: 490-491. https://doi.org/10.1038/nm0509-490
|
| [14] | Erenpreisa J, Giuliani A (2020) Resolution of Complex Issues in Genome Regulation and Cancer Requires Non-Linear and Network-Based Thermodynamics. Int J Mol Sci 21. https://doi.org/10.3390/ijms21010240 |
| [15] | Erenpreisa J, Salmina K, Anatskaya O, et al. (2020) Paradoxes of cancer: Survival at the brink. Semin Cancer Biol . https://doi.org/10.1016/j.semcancer.2020.12.009 |
| [16] | Krebs ET (1947) CANCER AND THE EMBRYONAL HYPOTHESIS. Calif Med 66: 270-271. |
| [17] |
Saitoh M (2018) Involvement of partial EMT in cancer progression. J Biochem 164: 257-264. https://doi.org/10.1093/jb/mvy047
|
| [18] |
Saitoh M (2018) JB special review-cellular plasticity in epithelial homeostasis and diseases: Involvement of partial EMT in cancer progression. J Biochem 164: 257-264. https://doi.org/10.1093/jb/mvy047
|
| [19] |
Mukund K, Syulyukina N, Ramamoorthy S, et al. (2020) Right and left-sided colon cancers-specificity of molecular mechanisms in tumorigenesis and progression. BMC Cancer 20. https://doi.org/10.1186/s12885-020-06784-7
|
| [20] |
Wessely A, Steeb T, Berking C, et al. (2021) How neural crest transcription factors contribute to melanoma heterogeneity, cellular plasticity, and treatment resistance. Int J Mol Sci 22. https://doi.org/10.3390/ijms22115761
|
| [21] |
Saravanan KM, Kannan M, Meera P, et al. (2022) E3 ligases: a potential multi-drug target for different types of cancers and neurological disorders. Future Med Chem 14: 187-201. https://doi.org/10.4155/fmc-2021-0157
|
| [22] |
Ben-Porath I, Thomson MW, Carey VJ, et al. (2008) An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat Genet 40: 499-507. https://doi.org/10.1038/ng.127
|
| [23] |
Viswanathan A, Musa A, Murugesan A, et al. (2019) Battling Glioblastoma: A Novel Tyrosine Kinase Inhibitor with Multi-Dimensional Anti-Tumor Effect (Running Title: Cancer Cells Death Signalling Activation). Cells 8. https://doi.org/10.3390/cells8121624
|
| [24] | Doan P, Nguyen P, Murugesan A, et al. (2021) Targeting orphan g protein-coupled receptor 17 with t0 ligand impairs glioblastoma growth. Cancers (Basel) 13. https://doi.org/10.3390/cancers13153773 |
| [25] |
Cofre J, Abdelhay E (2017) Cancer Is to Embryology as Mutation Is to Genetics: Hypothesis of the Cancer as Embryological Phenomenon. ScientificWorldJournal 2017: 3578090. https://doi.org/10.1155/2017/3578090
|
| [26] |
Rountree CB, Mishra L, Willenbring H (2012) Stem cells in liver diseases and cancer: recent advances on the path to new therapies. Hepatology 55: 298-306. https://doi.org/10.1002/hep.24762
|
| [27] |
Qin S, Jiang J, Lu Y, et al. (2020) Emerging role of tumor cell plasticity in modifying therapeutic response. Signal Transduct Target Ther 5: 228. https://doi.org/10.1038/s41392-020-00313-5
|
| [28] | Pierce GB (1983) The cancer cell and its control by the embryo. Rous-Whipple Award lecture. Am J Pathol 113: 117. |
| [29] |
Liu J (2018) The dualistic origin of human tumors. Semin Cancer Biol 53: 1-16. https://doi.org/10.1016/j.semcancer.2018.07.004
|
| [30] |
Xu B, Konze KD, Jin J, et al. (2015) Targeting EZH2 and PRC2 dependence as novel anticancer therapy. Exp Hematol 43: 698-712. https://doi.org/10.1016/j.exphem.2015.05.001
|
| [31] |
Wang W, Qin J, Voruganti S, et al. (2015) Polycomb group (PcG) proteins and human cancers: multifaceted functions and therapeutic implications. Med Res Rev 35: 1220-1267. https://doi.org/10.1002/med.21358
|
| [32] |
Murray MJ, Lessey BA (1999) Embryo implantation and tumor metastasis: Common pathways of invasion and angiogenesis. Semin Reprod Endocrinol 17: 275-290. https://doi.org/10.1055/s-2007-1016235
|
| [33] |
Costanzo V, Bardelli A, Siena S, et al. (2018) Exploring the links between cancer and placenta development. Open Biol 8: 180081. https://doi.org/10.1098/rsob.180081
|
| [34] |
Turajlic S, Sottoriva A, Graham T, et al. (2019) Resolving genetic heterogeneity in cancer. Nat Rev Genet 20: 404-416. https://doi.org/10.1038/s41576-019-0114-6
|
| [35] | Wagner GP, Kshitiz, Dighe A, et al. (2021) The Coevolution of Placentation and Cancer. Annu Rev Anim Biosci 10. https://doi.org/10.1146/annurev-animal-020420-031544 |
| [36] |
Fukuda MN, Sugihara K (2008) An integrated view of L-selectin and trophinin function in human embryo implantation. J Obstet Gynaecol Res 34: 129-136. https://doi.org/10.1111/j.1447-0756.2008.00776.x
|
| [37] |
Ayala GE, Dai H, Li R, et al. (2006) Bystin in perineural invasion of prostate cancer. Prostate 66: 266-272. https://doi.org/10.1002/pros.20323
|
| [38] |
Aplin JD, Ruane PT (2017) Embryo–epithelium interactions during implantation at a glance. J Cell Sci 130: 15-22. https://doi.org/10.1242/jcs.175943
|
| [39] |
Harada O, Suga T, Suzuki T, et al. (2007) The role of trophinin, an adhesion molecule unique to human trophoblasts, in progression of colorectal cancer. Int J Cancer 121: 1072-1078. https://doi.org/10.1002/ijc.22821
|
| [40] |
Sims GP, Rowe DC, Rietdijk ST, et al. (2009) HMGB1 and RAGE in inflammation and cancer. Annu Rev Immunol 28: 367-388. https://doi.org/10.1146/annurev.immunol.021908.132603
|
| [41] |
Ibrahim ZA, Armour CL, Phipps S, et al. (2013) RAGE and TLRs: relatives, friends or neighbours?. Mol Immunol 56: 739-744. https://doi.org/10.1016/j.molimm.2013.07.008
|
| [42] |
Cui T, Zhang W, Li S, et al. (2019) Oxidative Stress–Induced HMGB1 Release from Melanocytes: A Paracrine Mechanism Underlying the Cutaneous Inflammation in Vitiligo. J Invest Dermatol 139: 2174-2184.e4. https://doi.org/10.1016/j.jid.2019.03.1148
|
| [43] |
Rapoport BL, Steel HC, Theron AJ, et al. (2020) High Mobility Group Box 1 in Human Cancer. Cells 9. https://doi.org/10.3390/cells9071664
|
| [44] | Zhang H, Li J, Saravanan KM, et al. (2021) An Integrated Deep Learning and Molecular Dynamics Simulation-Based Screening Pipeline Identifies Inhibitors of a New Cancer Drug Target TIPE2. Front Pharmacol 12. https://doi.org/10.3389/fphar.2021.772296 |
| [45] |
Saravanan KM, Zhang H, Zhang H, et al. (2020) On the Conformational Dynamics of β-Amyloid Forming Peptides: A Computational Perspective. Front Bioeng Biotechnol 8. https://doi.org/10.3389/fbioe.2020.00532
|
| [46] |
Viswanathan A, Kute D, Musa A, et al. (2019) 2-(2-(2,4-dioxopentan-3-ylidene)hydrazineyl)benzonitrile as novel inhibitor of receptor tyrosine kinase and PI3K/AKT/mTOR signaling pathway in glioblastoma. Eur J Med Chem 166: 291-303. https://doi.org/10.1016/j.ejmech.2019.01.021
|
| [47] |
Imashiro C, Azuma T, Itai S, et al. (2021) Travelling ultrasound promotes vasculogenesis of three-dimensional-monocultured human umbilical vein endothelial cells. Biotechnol Bioeng 118: 3760-3769. https://doi.org/10.1002/bit.27852
|
| [48] | Srimathi Devi J, Haripriya D, Arul S, et al. (2021) Evaluation of anti-cancer effect of zerumbone and cisplatin on N-nitrosodiethylamine induced hepatic cancer in freshwater fish (Danio rerio). Nat Prod Res 1–5. https://doi.org/10.1080/14786419.2021.2012672 |
| [49] | Saravanan KM, Ponnuraj K (2018) Sequence and structural analysis of fibronectin-binding protein reveals importance of multiple intrinsic disordered tandem repeats. J Mol Recognit : e2768. https://doi.org/10.1002/jmr.2768 |
| [50] |
George EL, Georges-Labouesse EN, Patel-King RS, et al. (1993) Defects in mesoderm, neural tube and vascular development in mouse embryos lacking fibronectin. Development 119: 1079-1091. https://doi.org/10.1242/dev.119.4.1079
|
| [51] | Snow CJ, Goody M, Kelly MW, et al. (2008) Time-lapse analysis and mathematical characterization elucidate novel mechanisms underlying muscle morphogenesis. PLoS Genet 4. https://doi.org/10.1371/journal.pgen.1000219 |
| [52] | Murphy PA, Begum S, Hynes RO (2015) Tumor angiogenesis in the absence of fibronectin or its cognate integrin receptors. PLoS One 10. https://doi.org/10.1371/journal.pone.0120872 |
| [53] | Galmiche A, Rak J, Roumenina LT, et al. (2022) Coagulome and the tumor microenvironment: an actionable interplay. Trends in Cancer . https://doi.org/10.1016/j.trecan.2021.12.008 |
| [54] |
Nasser NJ, Fox J, Agbarya A (2020) Potential mechanisms of cancer-related hypercoagulability. Cancers (Basel) 12. https://doi.org/10.3390/cancers12030566
|
| [55] | Yu G, Wen Q (2018) Expression of embryonic liver fodrin (ELF) and stem cell markers in CD13 liver cancer stem cells. Eur Rev Med Pharmacol Sci 22: 1653-1657. |
| [56] |
Baek HJ, Lim SC, Kitisin K, et al. (2008) Hepatocellular cancer arises from loss of transforming growth factor beta signaling adaptor protein embryonic liver fodrin through abnormal angiogenesis. Hepatology 48: 1128-1137. https://doi.org/10.1002/hep.22460
|
| [57] |
Teng B, Huang C, Cheng CL, et al. (2020) Newly identified peptide hormone inhibits intestinal fat absorption and improves NAFLD through its receptor GPRC6A. J Hepatol 73: 383-393. https://doi.org/10.1016/j.jhep.2020.02.026
|
| [58] |
Bian J, Dannappel M, Wan C, et al. (2020) Transcriptional regulation of Wnt/β-catenin pathway in colorectal cancer. Cells 9: 2125. https://doi.org/10.3390/cells9092125
|
| [59] |
Davis-Marcisak EF, Deshpande A, Stein-O'Brien GL, et al. (2021) From bench to bedside: Single-cell analysis for cancer immunotherapy. Cancer Cell 39: 1062-1080. https://doi.org/10.1016/j.ccell.2021.07.004
|
| [60] |
Xie Y, Yao L, Yu X, et al. (2020) Action mechanisms and research methods of tRNA-derived small RNAs. Signal Transduct Target Ther 5: 1-9. https://doi.org/10.1038/s41392-020-00217-4
|
| [61] |
Postovit LM, Margaryan NV, Seftor EA, et al. (2008) Human embryonic stem cell microenvironment suppresses the tumorigenic phenotype of aggressive cancer cells. Proc Natl Acad Sci U S A 105: 4329-4334. https://doi.org/10.1073/pnas.0800467105
|
| [62] |
Dreesen O, Brivanlou AH (2007) Signaling pathways in cancer and embryonic stem cells. Stem Cell Rev 3: 7-17. https://doi.org/10.1007/s12015-007-0004-8
|
| [63] |
Saravanan KM, Palanivel S, Yli-Harja O, et al. (2018) Identification of novel GPR17-agonists by structural bioinformatics and signaling activation. Int J Biol Macromol 106: 901-907. https://doi.org/10.1016/j.ijbiomac.2017.08.088
|
| [64] |
Caruso S, O'Brien DR, Cleary SP, et al. (2021) Genetics of hepatocellular carcinoma: approaches to explore molecular diversity. Hepatology 73: 14-26. https://doi.org/10.1002/hep.31394
|
| [65] |
Gnanavel M, Murugesan A, Mani SK, et al. (2021) Identifying the mirna signature association with aging-related senescence in glioblastoma. Int J Mol Sci 22: 1-14. https://doi.org/10.3390/ijms22020517
|
| [66] |
Buhrmann C, Brockmueller A, Harsha C, et al. (2021) Evidence that tumor microenvironment initiates epithelial-to-mesenchymal transition and calebin a can suppress it in colorectal cancer cells. Front Pharmacol 12: 1689. https://doi.org/10.3389/fphar.2021.699842
|
| [67] | Zhang H, Shao X, Peng Y, et al. (2019) A novel machine learning based approach for iPS progenitor cell identification. PLoS Comput Biol . https://doi.org/10.1101/744920 |
| [68] |
Shetzer Y, Solomon H, Koifman G, et al. (2014) The paradigm of mutant p53-expressing cancer stem cells and drug resistance. Carcinogenesis 35: 1196-1208. https://doi.org/10.1093/carcin/bgu073
|
| [69] | Filippi L, Pini A, Cammalleri M, et al. (2021) β3-Adrenoceptor, a novel player in the round-trip from neonatal diseases to cancer: Suggestive clues from embryo. Med Res Rev : n/a. https://doi.org/10.1002/med.21874 |
| [70] |
Martinez NJ, Walhout AJM (2009) The interplay between transcription factors and microRNAs in genome-scale regulatory networks. Bioessays 31: 435-445. https://doi.org/10.1002/bies.200800212
|
| [71] |
Shi G, Jin Y (2010) Role of Oct4 in maintaining and regaining stem cell pluripotency. Stem Cell Res Ther 1: 1-9. https://doi.org/10.1186/scrt39
|
| [72] |
van den Berg DLC, Snoek T, Mullin NP, et al. (2010) An Oct4-centered protein interaction network in embryonic stem cells. Cell Stem Cell 6: 369-381. https://doi.org/10.1016/j.stem.2010.02.014
|
| [73] |
Innes KE, Byers TE (1999) Preeclampsia and breast cancer risk. Epidemiology 10: 722-732. https://doi.org/10.1097/00001648-199911000-00013
|
| [74] |
Mahendra G, Kumar S, Isayeva T, et al. (2005) Antiangiogenic cancer gene therapy by adeno-associated virus 2-mediated stable expression of the soluble FMS-like tyrosine kinase-1 receptor. Cancer Gene Ther 12: 26-34. https://doi.org/10.1038/sj.cgt.7700754
|
| [75] |
Bellamy L, Casas JP, Hingorani AD, et al. (2007) Pre-eclampsia and risk of cardiovascular disease and cancer in later life: Systematic review and meta-analysis. Br Med J 335: 974-977. https://doi.org/10.1136/bmj.39335.385301.BE
|
| [76] |
Wu P, Haththotuwa R, Kwok CS, et al. (2017) Preeclampsia and future cardiovascular health: a systematic review and meta-analysis. Circ Cardiovasc Qual Outcomes 10: e003497. https://doi.org/10.1161/CIRCOUTCOMES.116.003497
|
| [77] |
Fong GH, Rossant J, Gertsenstein M, et al. (1995) Role of the Flt-1 receptor tyrosine kinase in regulating the assembly of vascular endothelium. Nature 376: 66-70. https://doi.org/10.1038/376066a0
|
| [78] | Shibuya M, Yamaguchi S, Yamane A, et al. (1990) Nucleotide sequence and expression of a novel human receptor-type tyrosine kinase gene (flt) closely relatd to the fms family. Oncogene 5: 519-524. |
| [79] |
Kim JS, Kang EJ, Woo OH, et al. (2013) The relationship between preeclampsia, pregnancy-induced hypertension and maternal risk of breast cancer: A meta-analysis. Acta Oncol (Madr) 52: 1643-1648. https://doi.org/10.3109/0284186X.2012.750033
|
| [80] |
Calderon-Margalit R, Friedlander Y, Yanetz R, et al. (2009) Preeclampsia and subsequent risk of cancer: update from the Jerusalem Perinatal Study. Am J Obstet Gynecol 200: 63.e1-63.e5. https://doi.org/10.1016/j.ajog.2008.06.057
|
| [81] | Yoshitomi Y, Ikeda T, Saito H, et al. (2017) JunB regulates angiogenesis and neurovascular parallel alignment in mouse embryonic skin. J Cell Sci 130: 916-926. https://doi.org/10.1242/jcs.196303 |
| [82] |
Mihajlović AI, FitzHarris G (2018) Segregating chromosomes in the mammalian oocyte. Curr Biol 28: R895-R907. https://doi.org/10.1016/j.cub.2018.06.057
|
| [83] |
Wang J, Batourina E, Schneider K, et al. (2018) Polyploid superficial cells that maintain the urothelial barrier are produced via incomplete cytokinesis and endoreplication. Cell Rep 25: 464-477. https://doi.org/10.1016/j.celrep.2018.09.042
|
| [84] | Zhang J, Qiao Q, Xu H, et al. (2021) Human cell polyploidization: the good and the evil. Seminars in Cancer Biology . Elsevier. https://doi.org/10.1016/j.semcancer.2021.04.005 |
| [85] |
Vasudevan A, Schukken KM, Sausville EL, et al. (2021) Aneuploidy as a promoter and suppressor of malignant growth. Nat Rev Cancer 21: 89-103. https://doi.org/10.1038/s41568-020-00321-1
|
| [86] |
Zhu J, Tsai H-J, Gordon MR, et al. (2018) Cellular stress associated with aneuploidy. Dev Cell 44: 420-431. https://doi.org/10.1016/j.devcel.2018.02.002
|
| [87] |
Vázquez-Diez C, FitzHarris G (2018) Causes and consequences of chromosome segregation error in preimplantation embryos. Reproduction 155: R63-R76. https://doi.org/10.1530/REP-17-0569
|
| [88] |
Masset H, Tšuiko O, Vermeesch JR (2021) Genome-wide abnormalities in embryos: Origins and clinical consequences. Prenat Diagn 41: 554-563. https://doi.org/10.1002/pd.5895
|
| [89] | Sikora E, Czarnecka-Herok J, Bojko A, et al. (2020) Therapy-induced polyploidization and senescence: Coincidence or interconnection?. Seminars in Cancer Biology . Elsevier. https://doi.org/10.1016/j.semcancer.2020.11.015 |
| [90] |
Wang N, Hao F, Shi Y, et al. (2021) The Controversial Role of Polyploidy in Hepatocellular Carcinoma. Onco Targets Ther 14: 5335. https://doi.org/10.2147/OTT.S340435
|
| [91] |
Amend SR, Torga G, Lin K, et al. (2019) Polyploid giant cancer cells: Unrecognized actuators of tumorigenesis, metastasis, and resistance. Prostate 79: 1489-1497. https://doi.org/10.1002/pros.23877
|
| [92] | Was H, Borkowska A, Olszewska A, et al. (2021) Polyploidy formation in cancer cells: How a Trojan horse is born. Seminars in Cancer Biology . Elsevier. https://doi.org/10.1016/j.semcancer.2021.03.003 |
| [93] | Demin S, Berdieva M, Goodkov A, et al. (2021) Cell-cell fusions and cell-in-cell phenomena in healthy cells and cancer: Lessons from protists and invertebrates. Seminars in Cancer Biology . Elsevier. https://doi.org/10.1016/j.semcancer.2021.03.005 |
| [94] |
Zack TI, Schumacher SE, Carter SL, et al. (2013) Pan-cancer patterns of somatic copy number alteration. Nat Genet 45: 1134-1140. https://doi.org/10.1038/ng.2760
|
| [95] |
Chen J, Niu N, Zhang J, et al. (2019) Polyploid giant cancer cells (PGCCs): the evil roots of cancer. Curr Cancer Drug Targets 19: 360-367. https://doi.org/10.2174/1568009618666180703154233
|
| [96] |
Wen Q, Goldenson B, Silver SJ, et al. (2012) Identification of regulators of polyploidization presents therapeutic targets for treatment of AMKL. Cell 150: 575-589. https://doi.org/10.1016/j.cell.2012.06.032
|
| [97] |
Vakifahmetoglu H, Olsson M, Zhivotovsky B (2008) Death through a tragedy: mitotic catastrophe. Cell Death Differ 15: 1153-1162. https://doi.org/10.1038/cdd.2008.47
|
| [98] |
Ganem NJ, Storchova Z, Pellman D (2007) Tetraploidy, aneuploidy and cancer. Curr Opin Genet Dev 17: 157-162. https://doi.org/10.1016/j.gde.2007.02.011
|
| [99] |
Ganem NJ, Pellman D (2007) Limiting the proliferation of polyploid cells. Cell 131: 437-440. https://doi.org/10.1016/j.cell.2007.10.024
|
| [100] |
Leikam C, Hufnagel AL, Otto C, et al. (2015) In vitro evidence for senescent multinucleated melanocytes as a source for tumor-initiating cells. Cell Death Dis 6: e1711-e1711. https://doi.org/10.1038/cddis.2015.71
|
| [101] | Zheng L, Dai H, Zhou M, et al. (2012) Polyploid cells rewire DNA damage response networks to overcome replication stress-induced barriers for tumour progression. Nat Commun 3: 1-12. https://doi.org/10.1038/ncomms1825 |
| [102] |
Thura M, Ye Z, Al-Aidaroos AQ, et al. (2021) PRL3 induces polyploid giant cancer cells eliminated by PRL3-zumab to reduce tumor relapse. Commun Biol 4: 923. https://doi.org/10.1038/s42003-021-02449-8
|
| [103] |
Liu J (2020) The “life code”: A theory that unifies the human life cycle and the origin of human tumors. Semin Cancer Biol 60: 380-397. https://doi.org/10.1016/j.semcancer.2019.09.005
|
| [104] |
Niu N, Yao J, Bast RC, et al. (2021) IL-6 promotes drug resistance through formation of polyploid giant cancer cells and stromal fibroblast reprogramming. Oncogenesis 10: 65. https://doi.org/10.1038/s41389-021-00349-4
|
| [105] |
Campbell AM, Zhang Z-Y (2014) Phosphatase of regenerating liver: a novel target for cancer therapy. Expert Opin Ther Targets 18: 555-569. https://doi.org/10.1517/14728222.2014.892926
|
| [106] |
Shankaranarayanan JS, Kanwar JR, AL-Juhaishi AJA, et al. (2016) Doxorubicin Conjugated to Immunomodulatory Anticancer Lactoferrin Displays Improved Cytotoxicity Overcoming Prostate Cancer Chemo resistance and Inhibits Tumour Development in TRAMP Mice. Sci Rep 6: 32062. https://doi.org/10.1038/srep32062
|
| [107] |
Niu N, Mercado-Uribe I, Liu J (2017) Dedifferentiation into blastomere-like cancer stem cells via formation of polyploid giant cancer cells. Oncogene 36: 4887-4900. https://doi.org/10.1038/onc.2017.72
|
| [108] |
Xiong S, Feng Y, Cheng L (2019) Cellular reprogramming as a therapeutic target in cancer. Trends Cell Biol 29: 623-634. https://doi.org/10.1016/j.tcb.2019.05.001
|
| [109] |
Labi V, Erlacher M (2015) How cell death shapes cancer. Cell Death Dis 6: e1675-e1675. https://doi.org/10.1038/cddis.2015.20
|
| [110] |
Zhang D, Yang X, Yang Z, et al. (2017) Daughter Cells and Erythroid Cells Budding from PGCCs and Their Clinicopathological Significances in Colorectal Cancer. J Cancer 8: 469-478. https://doi.org/10.7150/jca.17012
|
| [111] |
Zhang S, Mercado-Uribe I, Hanash S, et al. (2013) iTRAQ-based proteomic analysis of polyploid giant cancer cells and budding progeny cells reveals several distinct pathways for ovarian cancer development. PLoS One 8: e80120. https://doi.org/10.1371/journal.pone.0080120
|
| [112] |
Lv H, Shi Y, Zhang L, et al. (2014) Polyploid giant cancer cells with budding and the expression of cyclin E, S-phase kinase-associated protein 2, stathmin associated with the grading and metastasis in serous ovarian tumor. BMC Cancer 14: 576. https://doi.org/10.1186/1471-2407-14-576
|
| [113] |
Xu J, Hardin H, Zhang R, et al. (2016) Stage-Specific Embryonic Antigen-1 (SSEA-1) Expression in Thyroid Tissues. Endocr Pathol 27: 271-275. https://doi.org/10.1007/s12022-016-9448-1
|
| [114] |
Fleming H (2021) Opaque Polyploid Cells in Ishikawa Endometrial Cultures Are Capable of Forming Megamitochondria, Organelles Derived from the Adaptation of Fused Mitochondria Whose Capacity to Develop Gaseous Vacuoles Suggests CO2 Retention and Hypoxic Metabolism. Adv Biosci Biotechnol 12: 229-255. https://doi.org/10.4236/abb.2021.127015
|
| [115] | Liu J (2021) Giant cells: Linking McClintock's heredity to early embryogenesis and tumor origin throughout millennia of evolution on Earth. Seminars in Cancer Biology . Elsevier. https://doi.org/10.1016/j.semcancer.2021.06.007 |
| [116] | Liu J, Niu N, Li X, et al. (2021) The life cycle of polyploid giant cancer cells and dormancy in cancer: Opportunities for novel therapeutic interventions. Semin Cancer Biol . https://doi.org/10.1016/j.semcancer.2021.10.005 |
| [117] | Li Z, Zheng M, Zhang H, et al. (2021) Arsenic Trioxide Promotes Tumor Progression by Inducing the Formation of PGCCs and Embryonic Hemoglobin in Colon Cancer Cells. Front Oncol 4046. https://doi.org/10.3389/fonc.2021.720814 |
| [118] |
Jang YJ, Won JH, Back MJ, et al. (2015) Paraquat induces apoptosis through a mitochondria-dependent pathway in RAW264.7 cells. Biomol Ther 23: 407-413. https://doi.org/10.4062/biomolther.2015.075
|
| [119] |
Smith DG, Sturmey RG (2013) Parallels between embryo and cancer cell metabolism. Biochem Soc Trans 41: 664-669. https://doi.org/10.1042/BST20120352
|
| [120] |
Christensen DR, Calder PC, Houghton FD (2015) GLUT3 and PKM2 regulate OCT4 expression and support the hypoxic culture of human embryonic stem cells. Sci Rep 5. https://doi.org/10.1038/srep17500
|
| [121] | Dang CV (2013) MYC, metabolism, cell growth, and tumorigenesis. Cold Spring Harb Perspect Med 3. https://doi.org/10.1101/cshperspect.a014217 |
| [122] |
Varlakhanova NV, Cotterman RF, deVries WN, et al. (2010) Myc maintains embryonic stem cell pluripotency and self-renewal. Differentiation 80: 9-19. https://doi.org/10.1016/j.diff.2010.05.001
|
| [123] |
Gabryelska A, Karuga FF, Szmyd B, et al. (2020) HIF-1α as a Mediator of Insulin Resistance, T2DM, and Its Complications: Potential Links With Obstructive Sleep Apnea. Front Physiol 11. https://doi.org/10.3389/fphys.2020.01035
|
| [124] | Yu M, Lun J, Zhang H, et al. (2021) The non-canonical functions of HIF prolyl hydroxylases and their dual roles in cancer. Int J Biochem Cell Biol 135. https://doi.org/10.1016/j.biocel.2021.105982 |
| [125] |
Kim J, Hong J, Lee J, et al. (2021) Recent advances in tumor microenvironment-targeted nanomedicine delivery approaches to overcome limitations of immune checkpoint blockade-based immunotherapy. J Control Release 332: 109-126. https://doi.org/10.1016/j.jconrel.2021.02.002
|
| [126] |
Parsons MJ, Tammela T, Dow LE (2021) WNT as a driver and dependency in cancer. Cancer Discov 11: 2413-2429. https://doi.org/10.1158/2159-8290.CD-21-0190
|
| [127] | Wang X, Wang Q (2018) Alpha-fetoprotein and hepatocellular carcinoma immunity. Can J Gastroenterol Hepatol 2018. https://doi.org/10.1155/2018/9049252 |
| [128] |
Vujanovic L, Stahl EC, Pardee AD, et al. (2017) Tumor-derived a-fetoprotein directly drives human natural killer-cell activation and subsequent cell death. Cancer Immunol Res 5: 493-502. https://doi.org/10.1158/2326-6066.CIR-16-0216
|
| [129] |
Samet JM, Avila-Tang E, Boffetta P, et al. (2009) Lung cancer in never smokers: Clinical epidemiology and environmental risk factors. Clin Cancer Res 15: 5626-5645. https://doi.org/10.1158/1078-0432.CCR-09-0376
|
| [130] |
Boogerd LSF, Handgraaf HJM, Lam HD, et al. (2017) Laparoscopic detection and resection of occult liver tumors of multiple cancer types using real-time near-infrared fluorescence guidance. Surg Endosc 31: 952-961. https://doi.org/10.1007/s00464-016-5007-6
|
| [131] |
Samanta S, Guru S, Elaimy AL, et al. (2018) IMP3 Stabilization of WNT5B mRNA Facilitates TAZ Activation in Breast Cancer. Cell Rep 23: 2559-2567. https://doi.org/10.1016/j.celrep.2018.04.113
|
| [132] | Beilerli A, Gareev I, Beylerli O, et al. (2021) Circular RNAs as biomarkers and therapeutic targets in cancer. Seminars in Cancer Biology . Elsevier. |
| [133] | Zhu T, Tu SZ, Yang YL, et al. (2019) Induced effects of resveratrol on apoptosis and autophagy in human breast cancer MDA-MB231 cells. Chinese Pharmacol Bull 35: 839-843. https://doi.org/10.1016/j.semcancer.2020.12.026 |
| [134] |
Wu Y, Wang F, Albert Reece E, et al. (2015) Curcumin ameliorates high glucose-induced neural tube defects by suppressing cellular stress and apoptosis. Am J Obstet Gynecol 212: 802.e1-802.e8. https://doi.org/10.1016/j.ajog.2015.01.017
|
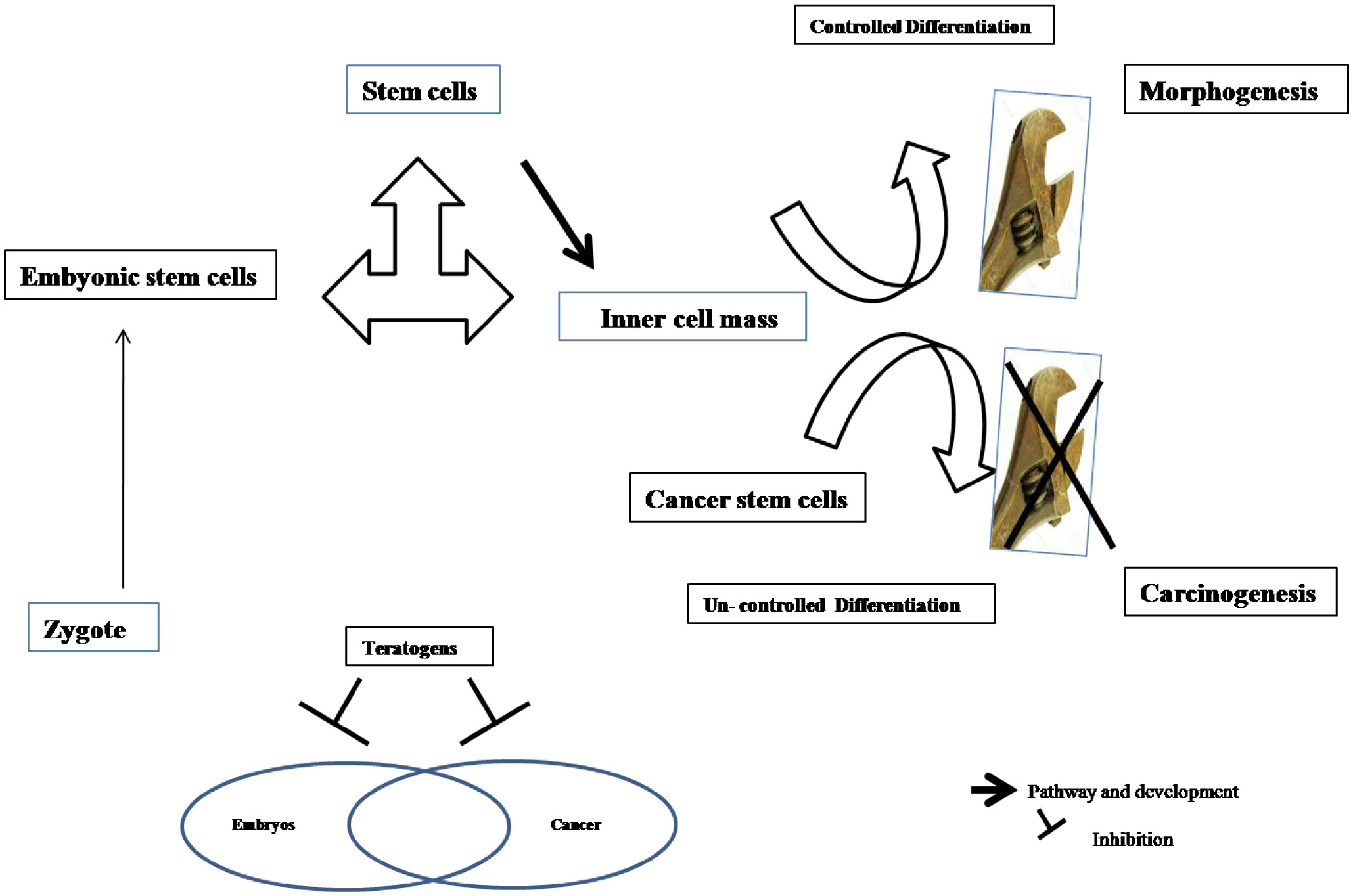
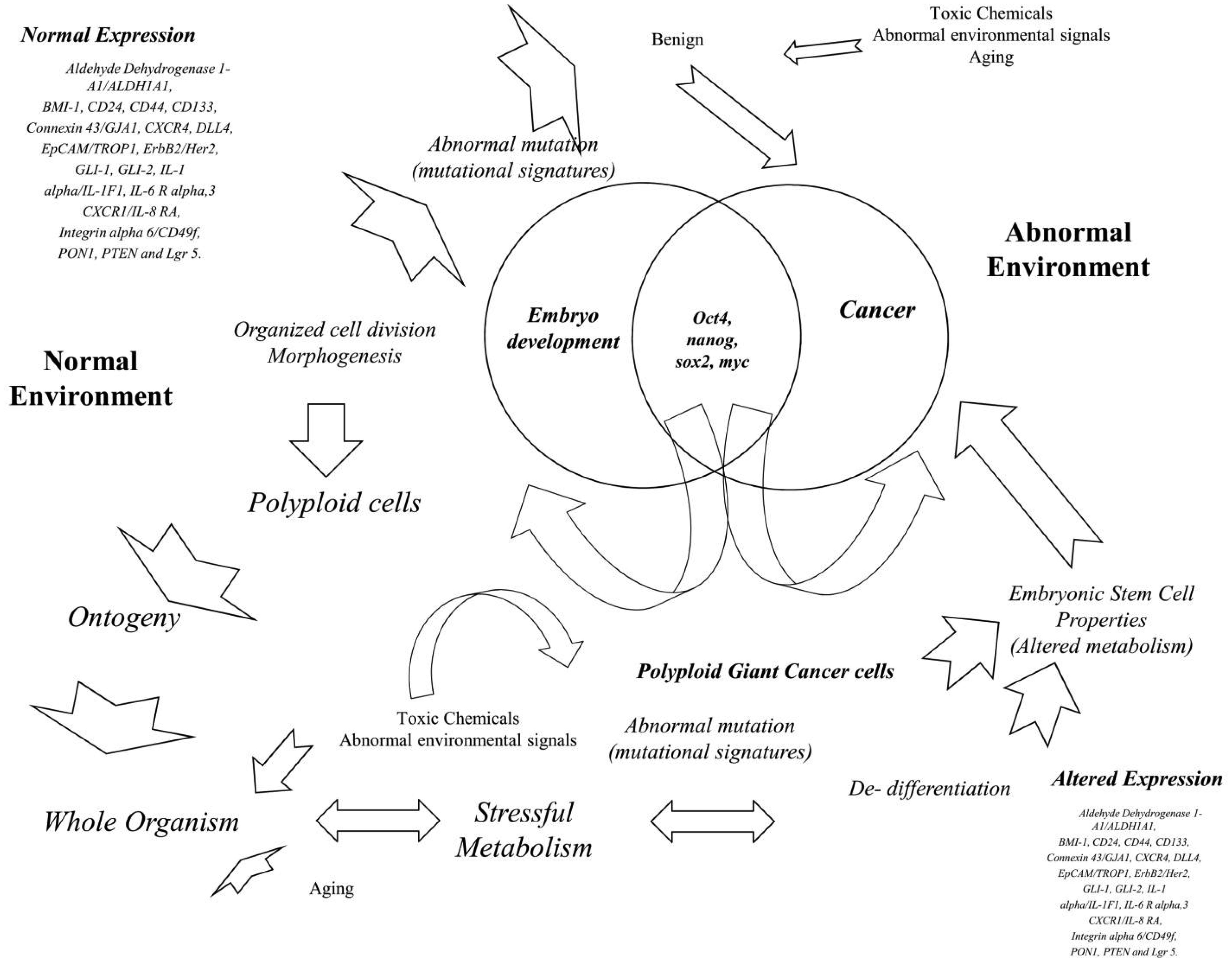
Figures(2)

Kishore Kumar Meenakshi Sundaram, Giridharan Bupesh, Konda Mani Saravanan. Instrumentals behind embryo and cancer: a platform for prospective future in cancer research[J]. AIMS Molecular Science, 2022, 9(1): 25-45. doi: 10.3934/molsci.2022002







 DownLoad:
DownLoad: