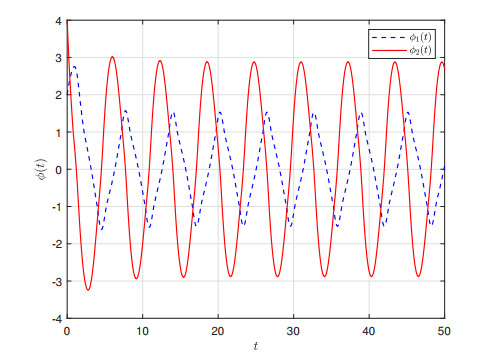
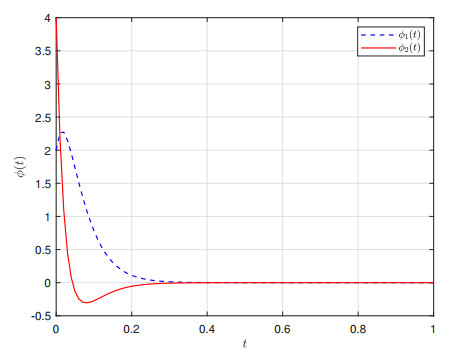
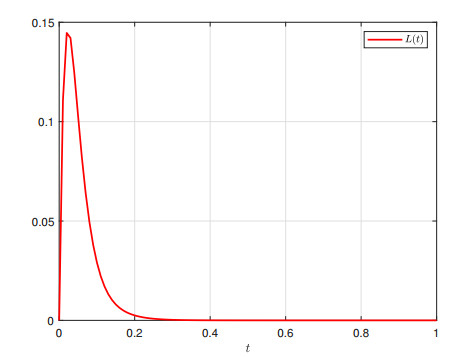
This paper investigates $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ control for memristive neural networks (MNNs) with a non-necessarily differentiable time-varying delay. The objective is to design an output-feedback controller to ensure the $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ stability of the considered MNN. A criterion on the $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ stability is proposed using a Lyapunov functional, the Bessel-Legendre inequality, and the convex combination inequality. Then, a linear matrix inequalities-based design scheme for the required output-feedback controller is developed by decoupling nonlinear terms. Finally, two examples are presented to verify the proposed $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ stability criterion and design method.
Citation: Jingya Wang, Ye Zhu. $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ control for memristive NNs with non-necessarily differentiable time-varying delay[J]. Mathematical Biosciences and Engineering, 2023, 20(7): 13182-13199. doi: 10.3934/mbe.2023588
This paper investigates $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ control for memristive neural networks (MNNs) with a non-necessarily differentiable time-varying delay. The objective is to design an output-feedback controller to ensure the $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ stability of the considered MNN. A criterion on the $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ stability is proposed using a Lyapunov functional, the Bessel-Legendre inequality, and the convex combination inequality. Then, a linear matrix inequalities-based design scheme for the required output-feedback controller is developed by decoupling nonlinear terms. Finally, two examples are presented to verify the proposed $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ stability criterion and design method.

| [1] |
Z. Zhang, X. Liu, D. Zhou, C. Lin, J. Chen, H. Wang, Finite-time stabilizability and instabilizability for complex-valued memristive neural networks with time delays, IEEE Trans. Syst. Man Cybern. Syst., 48 (2017), 2371–2382. https://doi.org/10.1109/TSMC.2017.2754508 doi: 10.1109/TSMC.2017.2754508

|
| [2] |
A. Wu, Z. Zeng, Exponential stabilization of memristive neural networks with time delays, IEEE Trans. Neural Networks Learn. Syst., 23 (2012), 1919–1929. https://doi.org/10.1109/TNNLS.2012.2219554 doi: 10.1109/TNNLS.2012.2219554
|
| [3] |
R. Li, X. Gao, J. Cao, Quasi-state estimation and quasi-synchronization control of quaternion-valued fractional-order fuzzy memristive neural networks: Vector ordering approach, Appl. Math. Comput., 362 (2019), 124572. https://doi.org/10.1016/j.amc.2019.124572 doi: 10.1016/j.amc.2019.124572
|
| [4] |
J. Zhang, X. Ma, Y. Li, Q. Gan, C. Wang, Synchronization in fixed/preassigned-time of delayed fully quaternion-valued memristive neural networks via non-separation method, Commun. Nonlinear Sci. Numer. Simul., 113 (2022), 106581. https://doi.org/10.1016/j.cnsns.2022.106581 doi: 10.1016/j.cnsns.2022.106581
|
| [5] |
N. Yang, Y. Yu, S. Zhong, X. Wang, K. Shi, J. Cai, Exponential synchronization of stochastic delayed memristive neural networks via a novel hybrid control, Neural Networks, 131 (2020), 242–250. https://doi.org/10.1016/j.neunet.2020.07.034 doi: 10.1016/j.neunet.2020.07.034
|
| [6] | Z. Yan, X. Huang, J. Cao, Variable-sampling-period dependent global stabilization of delayed memristive neural networks based on refined switching event-triggered control, Sci. China Inf. Sci., 63 (2020), 212201. |
| [7] | X. Wu, S. Liu, H. Wang, Asymptotic stability and synchronization of fractional delayed memristive neural networks with algebraic constraints, Commun. Nonlinear Sci. Numer. Simul., 114 (2022), 106694. |
| [8] |
Z. Yan, X. Huang, Y. Fan, J. Xia, H. Shen, Threshold-function-dependent quasi-synchronization of delayed memristive neural networks via hybrid event-triggered control, IEEE Trans. Syst. Man Cybern. Syst., 51 (2021), 6712–6722. https://doi.org/10.1109/TSMC.2020.2964605 doi: 10.1109/TSMC.2020.2964605
|
| [9] | S. Murugesan, Y. C. Liu, Finite-time resilient control for networked control systems with multiple cyber-attacks: Memory/adaptive event-triggered scheme, Int. J. Adapt. Control Signal Process., 36 (2022), 901–925. |
| [10] |
W. Tai, D. Gao, A. Zhao, J. Zhou, X. Wang, Weight learning for $\mathcal{H}_{\infty}$ stabilization of uncertain switched neural networks with external disturbance and reaction-diffusion, Int. J. Adapt. Control Signal Process., 37 (2023), 1015–1029. https://doi.org/10.1002/acs.3558 doi: 10.1002/acs.3558
|
| [11] |
M. Sathishkumar, R. Sakthivel, F. Alzahrani, B. Kaviarasan, Y. Ren, Mixed $\mathcal{H}_{\infty}$ and passivity-based resilient controller for nonhomogeneous Markov jump systems, Nonlinear Anal. Hybrid Syst., 31 (2019), 86–99. https://doi.org/10.1016/j.mattod.2019.06.003 doi: 10.1016/j.mattod.2019.06.003
|
| [12] |
J. Zhou, J. Dong, S. Xu, Asynchronous dissipative control of discrete-time fuzzy Markov jump systems with dynamic state and input quantization, IEEE Trans. Fuzzy Syst., 2023 (2023), forthcoming. https://doi.org/10.1109/TFUZZ.2023.3271348 doi: 10.1109/TFUZZ.2023.3271348
|
| [13] |
X. Meng, G. Zhang, Q. Zhang, Robust adaptive neural network integrated fault-tolerant control for underactuated surface vessels with finite-time convergence and event-triggered inputs, Math. Biosci. Eng., 20 (2022), 2131–2156. https://doi.org/10.3934/mbe.2023099 doi: 10.3934/mbe.2023099
|
| [14] |
R. Saravanakumar, Y. Hoon Joo, Network-based robust exponential fuzzy control for uncertain systems, Math. Methods Appl. Sci., 2023 (2023), forthcoming. https://doi.org/10.1002/mma.8943 doi: 10.1002/mma.8943
|
| [15] |
Y. Cao, N. Liu, C. Zhang, T. Zhang, Z.-F. Luo, Synchronization of multiple reaction–diffusion memristive neural networks with known or unknown parameters and switching topologies, Knowl. Based Syst., 254 (2022), 109595. https://doi.org/10.1016/j.knosys.2022.109595 doi: 10.1016/j.knosys.2022.109595
|
| [16] | I. Ghous, J. Lu, Z. Duan, $\mathcal{H}_{\infty}$ stabilization problem for memristive neural networks with time-varying delays, Inf. Sci., 607 (2022), 27–43. |
| [17] |
Z. Yan, D. Zuo, T. Guo, J. Zhou, Quantized $\mathcal{H}_{\infty}$ stabilization for delayed memristive neural networks, Neural Comput. Appl., 2023 (2023), forthcoming. https://doi.org/10.1007/s00521-023-08510-3 doi: 10.1007/s00521-023-08510-3
|
| [18] |
W. Tai, X. Li, J. Zhou, S. Arik, Asynchronous dissipative stabilization for stochastic Markov-switching neural networks with completely-and incompletely-known transition rates, Neural Networks, 161 (2023), 55–64. https://doi.org/10.1016/j.neunet.2023.01.039 doi: 10.1016/j.neunet.2023.01.039
|
| [19] |
C. Deng, D. Zhang, G. Feng, Resilient practical cooperative output regulation for mass with unknown switching exosystem dynamics under DoS attacks, Automatica, 139 (2022), 110172. https://doi.org/10.1016/j.automatica.2022.110172 doi: 10.1016/j.automatica.2022.110172
|
| [20] | S. Dong, L. Liu, G. Feng, M. Liu, Z. G. Wu, R. Zheng, Cooperative output regulation quadratic control for discrete-time heterogeneous multiagent Markov jump systems, IEEE Trans. Cybern., 52 (2022), 9882–9892. |
| [21] | J. Gu, H. Wang, W. Li, Output-feedback stabilization for stochastic nonlinear systems with Markovian switching and time-varying powers, Math. Biosci. Eng., 19 (2022), 11 071–11 085. |
| [22] |
T. Yu, L. Liu, Y. J. Liu, Observer-based adaptive fuzzy output feedback control for functional constraint systems with dead-zone input, Math. Biosci. Eng., 20 (2023), 2628–2650. https://doi.org/10.3934/mbe.2023123 doi: 10.3934/mbe.2023123
|
| [23] | Z. Yan, C. Sang, M. Fang, J. Zhou, Energy-to-peak consensus for multi-agent systems with stochastic disturbances and Markovian switching topologies, Trans. Inst. Measure. Control, 40 (2018), 4358–4368. |
| [24] |
J. Zhou, D. Xu, W. Tai, C. K. Ahn, Switched event-triggered $\mathcal{H}_{\infty}$ security control for networked systems vulnerable to aperiodic DoS attacks, IEEE Trans. Network Sci. Eng., 2023 (2023), forthcoming. https://doi.org/10.1109/TNSE.2023.3243095 doi: 10.1109/TNSE.2023.3243095
|
| [25] |
Y. Liu, J. Li, F. Fang, J. H. Park, Further studies on sampled-data consensus of multi-agent systems with communication delays, IEEE Trans. Signal Inf. Process. Networks, 8 (2022), 920–931. https://doi.org/10.1109/TSIPN.2022.3220153 doi: 10.1109/TSIPN.2022.3220153
|
| [26] |
E. Arslan, G. Narayanan, M. S. Ali, S. Arik, S. Saroha, Controller design for finite-time and fixed-time stabilization of fractional-order memristive complex-valued bam neural networks with uncertain parameters and time-varying delays, Neural Networks, 130 (2020), 60–74. https://doi.org/10.1016/j.neunet.2020.06.021 doi: 10.1016/j.neunet.2020.06.021
|
| [27] |
R. Saravanakumar, G. Rajchakit, M. S. Ali, Z. Xiang, Y. H. Joo, Robust extended dissipativity criteria for discrete-time uncertain neural networks with time-varying delays, Neural Comput. Appl., 30 (2018), 3893–3904. https://doi.org/10.1007/s00521-017-2974-z doi: 10.1007/s00521-017-2974-z
|
| [28] |
L. He, W. Wu, J. Zhou, G. Yao, Input-to-state stable synchronization for delayed Lurie systems via sampled-data control, Discrete Contin. Dyn. Syst., 28 (2023), 1553–1570. https://doi.org/10.3934/dcdsb.2022135 doi: 10.3934/dcdsb.2022135
|
| [29] | X. Huang, J. Jia, Y. Fan, Z. Wang, J. Xia, Interval matrix method based synchronization criteria for fractional-order memristive neural networks with multiple time-varying delays, J. Franklin Inst., 357 (2020), 1707–1733. |
| [30] |
L. Yao, Z. Wang, X. Huang, Y. Li, Q. Ma, H. Shen, Stochastic sampled-data exponential synchronization of Markovian jump neural networks with time-varying delays, IEEE Trans. Neural Networks Learn. Syst., 34 (2023), 909–920. https://doi.org/10.1109/TNNLS.2021.3103958 doi: 10.1109/TNNLS.2021.3103958
|
| [31] |
R. Vadivel, P. Hammachukiattikul, N. Gunasekaran, R. Saravanakumar, H. Dutta, Strict dissipativity synchronization for delayed static neural networks: An event-triggered scheme, Chaos Solitons Fractals, 150 (2021), 111212. https://doi.org/10.1016/j.chaos.2021.111212 doi: 10.1016/j.chaos.2021.111212
|
| [32] | Y. Ni, Z. Wang, Y. Fan, X. Huang, H. Shen, Memory-based event-triggered control for global synchronization of chaotic Lur'e systems and its application, IEEE Trans. Syst. Man Cybern. Syst., 53 (2023), 1920–1931. |
| [33] |
G. Zhang, Z. Zeng, D. Ning, Novel results on synchronization for a class of switched inertial neural networks with distributed delays, Inf. Sci., 511 (2020), 114–126. https://doi.org/10.1016/j.ins.2019.09.048 doi: 10.1016/j.ins.2019.09.048
|
| [34] |
Z. Wang, S. Ding, Z. Huang, H. Zhang, Exponential stability and stabilization of delayed memristive neural networks based on quadratic convex combination method, IEEE Trans. Neural Networks Learn. Syst., 27 (2015), 2337–2350. https://doi.org/10.1109/TNNLS.2015.2485259 doi: 10.1109/TNNLS.2015.2485259
|
| [35] |
X. Yang, D. W. Ho, Synchronization of delayed memristive neural networks: robust analysis approach, IEEE Trans. Cybern., 46 (2015), 3377–3387. https://doi.org/10.1109/TCYB.2015.2505903 doi: 10.1109/TCYB.2015.2505903
|
| [36] |
A. Seuret, F. Gouaisbaut, Hierarchy of LMI conditions for the stability analysis of time-delay systems, Syst. Control Lett., 81 (2015), 1–7. https://doi.org/10.1016/j.sysconle.2015.03.007 doi: 10.1016/j.sysconle.2015.03.007
|
| [37] |
P. Park, J. W. Ko, C. Jeong, Reciprocally convex approach to stability of systems with time-varying delays, Automatica, 47 (2011), 235–238. https://doi.org/10.1016/j.automatica.2010.10.014 doi: 10.1016/j.automatica.2010.10.014
|
| [38] |
S. Xu, J. Lam, A survey of linear matrix inequality techniques in stability analysis of delay systems, Int. J. Syst. Sci., 39 (2008), 1095–1113. https://doi.org/10.1080/00207720802300370 doi: 10.1080/00207720802300370
|
| [39] | S. Boyd, L. El Ghaoui, E. Feron, V. Balakrishnan, Linear Matrix Inequalities in System and Control Theory, SIAM, 1994. |
| [40] |
J. Zhou, J. H. Park, Q. Ma, Non-fragile observer-based $\mathcal{H}_{\infty}$ control for stochastic time-delay systems, Appl. Math. Comput., 291 (2016), 69–83. https://doi.org/10.1016/j.amc.2016.06.024 doi: 10.1016/j.amc.2016.06.024
|
| [41] |
M. Solak, O. Faydasicok, S. Arik, A general framework for robust stability analysis of neural networks with discrete time delays, Neural Networks, 162 (2023), 186–198. https://doi.org/10.1016/j.neunet.2023.02.040 doi: 10.1016/j.neunet.2023.02.040
|
Figures(3) / Tables(1)

Jingya Wang, Ye Zhu. $ \mathcal{L}_{2}-\mathcal{L}_{\infty} $ control for memristive NNs with non-necessarily differentiable time-varying delay[J]. Mathematical Biosciences and Engineering, 2023, 20(7): 13182-13199. doi: 10.3934/mbe.2023588







 DownLoad:
DownLoad: