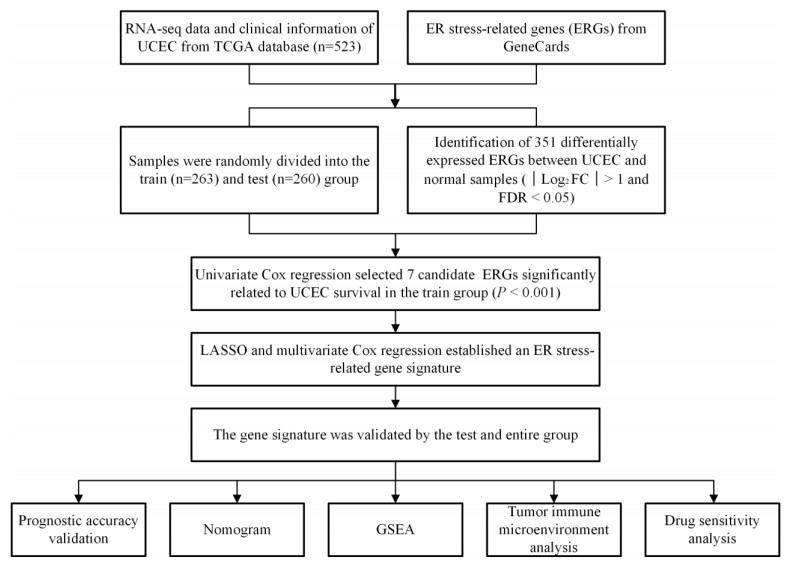
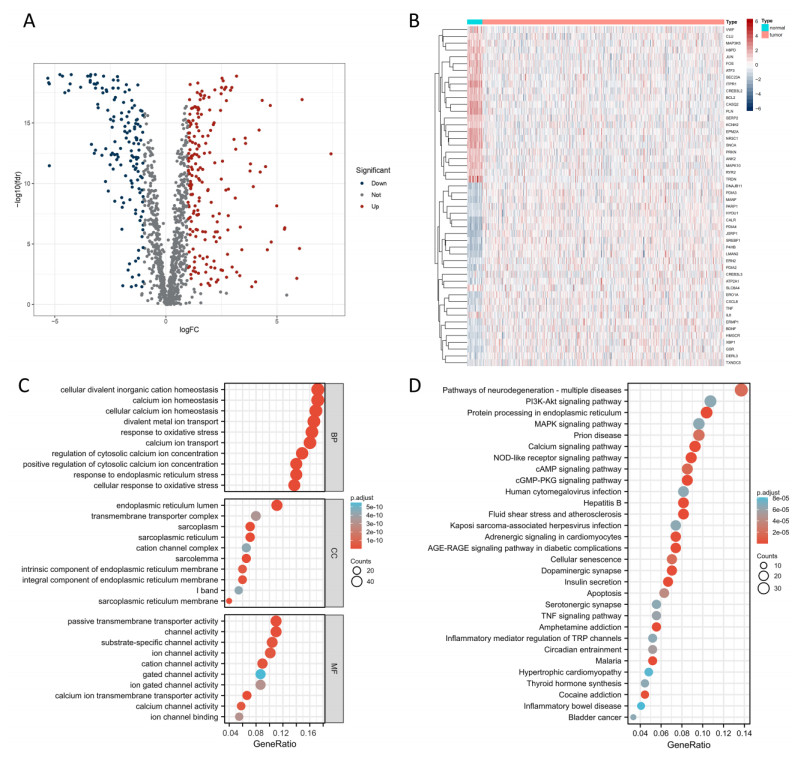
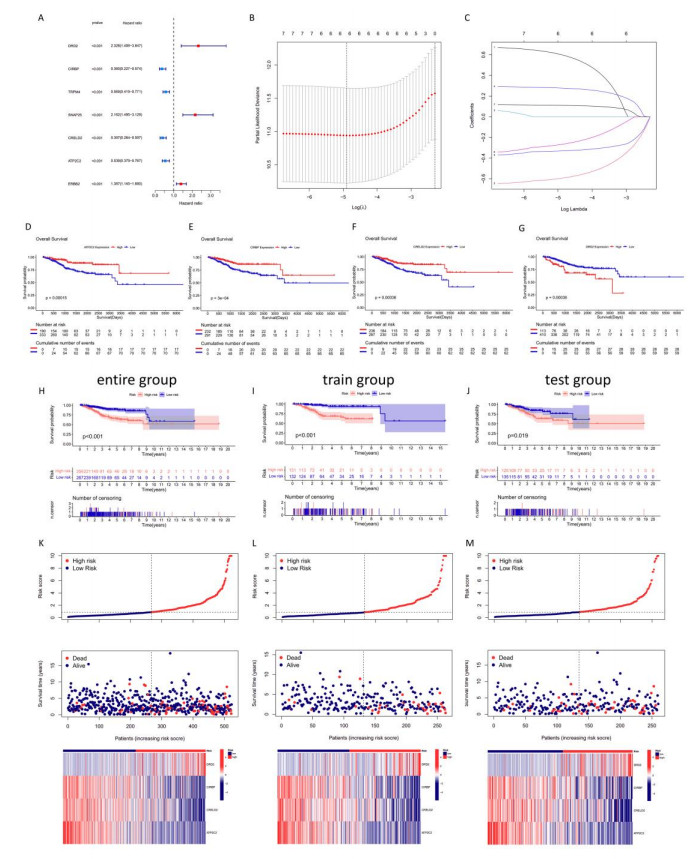
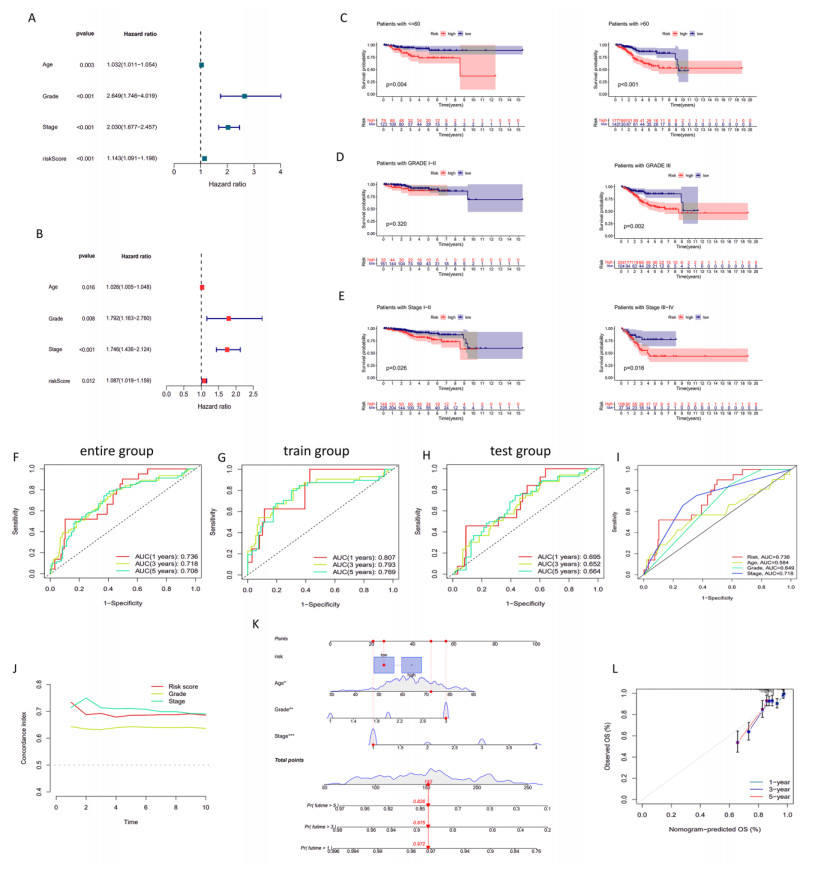
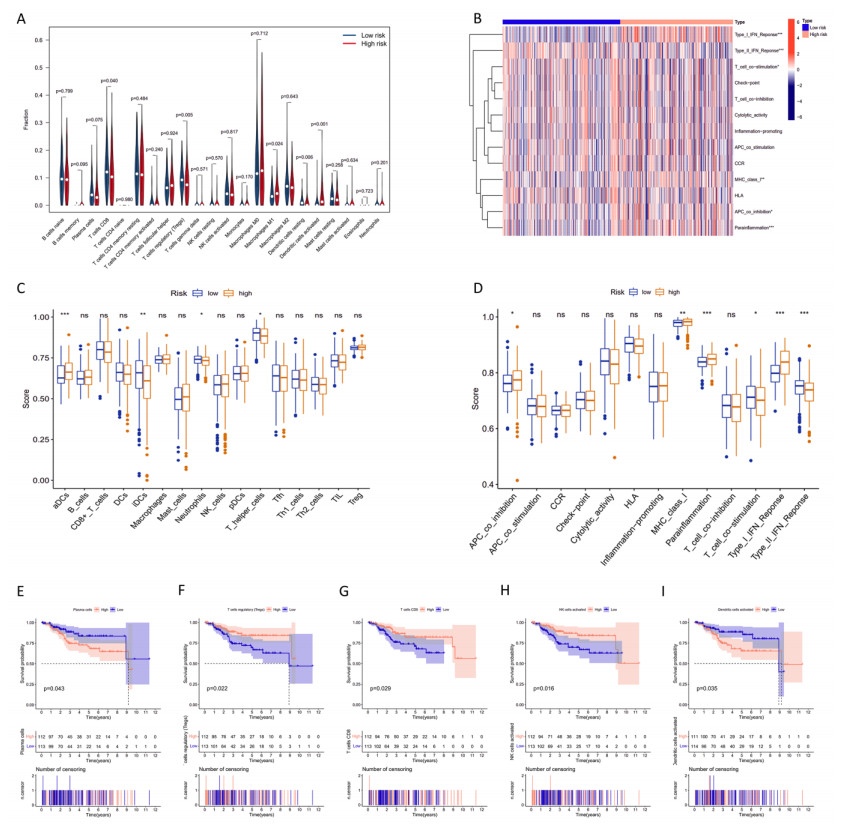
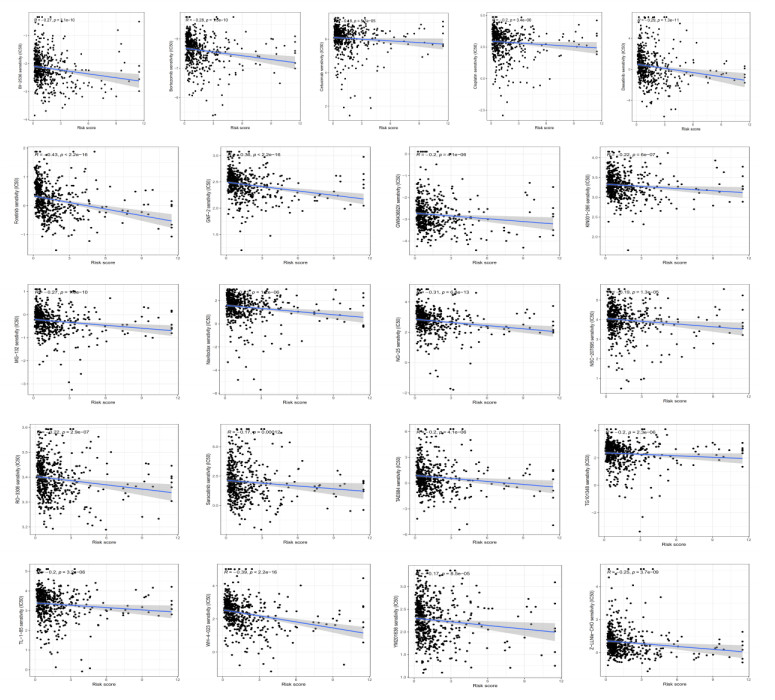
Uterine corpus endometrial cancer (UCEC) is the sixth most common female cancer worldwide, with an increasing incidence. Improving the prognosis of patients living with UCEC is a top priority. Endoplasmic reticulum (ER) stress has been reported to be involved in tumor malignant behaviors and therapy resistance, but its prognostic value in UCEC has been rarely investigated. The present study aimed to construct an ER stress-related gene signature for risk stratification and prognosis prediction in UCEC. The clinical and RNA sequencing data of 523 UCEC patients were extracted from TCGA database and were randomly assigned into a test group (n = 260) and training group (n = 263). An ER stress-related gene signature was established by LASSO and multivariate Cox regression in the training group and validated by Kaplan-Meier survival analysis, Receiver Operating Characteristic (ROC) curves and nomograms in the test group. Tumor immune microenvironment was analyzed by CIBERSORT algorithm and single-sample gene set enrichment analysis. R packages and the Connectivity Map database were used to screen the sensitive drugs. Four ERGs (ATP2C2, CIRBP, CRELD2 and DRD2) were selected to build the risk model. The high-risk group had significantly reduced overall survival (OS) (P < 0.05). The risk model had better prognostic accuracy than clinical factors. Tumor-infiltrating immune cells analysis depicted that CD8+ T cells and regulatory T cells were more abundant in the low-risk group, which may be related to better OS, while activated dendritic cells were active in the high-risk group and associated with unfavorable OS. Several kinds of drugs sensitive to the high-risk group were screened out. The present study constructed an ER stress-related gene signature, which has the potential to predict the prognosis of UCEC patients and have implications for UCEC treatment.
Citation: Pei Zhou, Caiyun Wu, Cong Ma, Ting Luo, Jing Yuan, Ping Zhou, Zhaolian Wei. Identification of an endoplasmic reticulum stress-related gene signature to predict prognosis and potential drugs of uterine corpus endometrial cancer[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 4018-4039. doi: 10.3934/mbe.2023188
Uterine corpus endometrial cancer (UCEC) is the sixth most common female cancer worldwide, with an increasing incidence. Improving the prognosis of patients living with UCEC is a top priority. Endoplasmic reticulum (ER) stress has been reported to be involved in tumor malignant behaviors and therapy resistance, but its prognostic value in UCEC has been rarely investigated. The present study aimed to construct an ER stress-related gene signature for risk stratification and prognosis prediction in UCEC. The clinical and RNA sequencing data of 523 UCEC patients were extracted from TCGA database and were randomly assigned into a test group (n = 260) and training group (n = 263). An ER stress-related gene signature was established by LASSO and multivariate Cox regression in the training group and validated by Kaplan-Meier survival analysis, Receiver Operating Characteristic (ROC) curves and nomograms in the test group. Tumor immune microenvironment was analyzed by CIBERSORT algorithm and single-sample gene set enrichment analysis. R packages and the Connectivity Map database were used to screen the sensitive drugs. Four ERGs (ATP2C2, CIRBP, CRELD2 and DRD2) were selected to build the risk model. The high-risk group had significantly reduced overall survival (OS) (P < 0.05). The risk model had better prognostic accuracy than clinical factors. Tumor-infiltrating immune cells analysis depicted that CD8+ T cells and regulatory T cells were more abundant in the low-risk group, which may be related to better OS, while activated dendritic cells were active in the high-risk group and associated with unfavorable OS. Several kinds of drugs sensitive to the high-risk group were screened out. The present study constructed an ER stress-related gene signature, which has the potential to predict the prognosis of UCEC patients and have implications for UCEC treatment.

| [1] |
H. Sung, J. Ferlay, R. L. Siegel, M. Laversanne, I. Soerjomataram, A. Jemal, et al., Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries, CA: Cancer J. Clin., 71 (2021), 209–249. https://doi.org/10.3322/caac.21660 doi: 10.3322/caac.21660

|
| [2] |
K. D. Miller, L. Nogueira, T. Devasia, A. B. Mariotto, K. R. Yabroff, A. Jemal, et al., Cancer treatment and survivorship statistics, 2022, CA: Cancer J. Clin., 72 (2022), 409–436. https://doi.org/10.3322/caac.21731 doi: 10.3322/caac.21731
|
| [3] |
M. Koskas, F. Amant, M. R. Mirza, C. L. Creutzberg, Cancer of the corpus uteri: 2021 update, Int. J. Gynaecol. Obstet., 155 (2021), 45–60. https://doi.org/10.1002/ijgo.13866 doi: 10.1002/ijgo.13866
|
| [4] |
J. R. Cubillos-Ruiz, S. E. Bettigole, L. H. Glimcher, Tumorigenic and immunosuppressive effects of endoplasmic reticulum stress in cancer, Cell, 168 (2017), 692–706. https://doi.org/10.1016/j.cell.2016.12.004 doi: 10.1016/j.cell.2016.12.004
|
| [5] |
X. Chen, J. R. Cubillos-Ruiz, Endoplasmic reticulum stress signals in the tumour and its microenvironment, Nat. Rev. Cancer, 21 (2021), 71–88. https://doi.org/10.1038/s41568-020-00312-2 doi: 10.1038/s41568-020-00312-2
|
| [6] |
Y. Hussain, H. Khan, T. Efferth, W. Alam, Regulation of endoplasmic reticulum stress by hesperetin: Focus on antitumor and cytoprotective effects, Phytomedicine, 100 (2022), 153985. https://doi.org/10.1016/j.phymed.2022.153985 doi: 10.1016/j.phymed.2022.153985
|
| [7] |
H. Urra, E. Dufey, T. Avril, E. Chevet, C. Hetz, Endoplasmic reticulum stress and the hallmarks of cancer, Trends Cancer, 2 (2016), 252–262. https://doi.org/10.1016/j.trecan.2016.03.007 doi: 10.1016/j.trecan.2016.03.007
|
| [8] |
W. M. Shi, Z. X. Chen, L. F. Li, H. Liu, R. Zhang, Q. L. Cheng, et al., Unravel the molecular mechanism of XBP1 in regulating the biology of cancer cells, J. Cancer, 10 (2019), 2035–2046. https://doi.org/10.7150/jca.29421 doi: 10.7150/jca.29421
|
| [9] |
J. Khaled, M. Kopsida, H. Lennernas, F. Heindryckx, Drug resistance and endoplasmic reticulum stress in hepatocellular carcinoma, Cells, 11 (2022), 632. https://doi.org/10.3390/cells11040632 doi: 10.3390/cells11040632
|
| [10] |
C. Salvagno, J. K. Mandula, P. C. Rodriguez, J. R. Cubillos-Ruiz, Decoding endoplasmic reticulum stress signals in cancer cells and antitumor immunity, Trends Cancer, 8 (2022), 930–943. https://doi.org/10.1016/j.trecan.2022.06.006 doi: 10.1016/j.trecan.2022.06.006
|
| [11] |
L. Sisinni, M. Pietrafesa, S. Lepore, F. Maddalena, V. Condelli, F. Esposito, et al., Endoplasmic reticulum stress and unfolded protein response in breast cancer: The balance between apoptosis and autophagy and iits role in drug resistance, Int. J. Mol. Sci., 20 (2019), 857. https://doi.org/10.3390/ijms20040857 doi: 10.3390/ijms20040857
|
| [12] |
M. Markouli, D. Strepkos, A. G. Papavassiliou, C. Piperi, Targeting of endoplasmic reticulum (ER) stress in gliomas, Pharmacol. Res., 157 (2020), 104823. https://doi.org/10.1016/j.phrs.2020.104823 doi: 10.1016/j.phrs.2020.104823
|
| [13] |
J. C. Wu, S. Qiao, Y. E. Xiang, M. Y. Cui, X. X. Yao, R. X. Lin, et al., Endoplasmic reticulum stress: Multiple regulatory roles in hepatocellular carcinoma, Biomed. Pharmacother., 142 (2021), 112005. https://doi.org/10.1016/j.biopha.2021.112005 doi: 10.1016/j.biopha.2021.112005
|
| [14] |
L. P. Wang, P. C. Fu, Y. Zhao, G. Wang, R. Yu, X. Wang, et al., Dissociation of NSC606985 induces atypical ER-stress and cell death in prostate cancer cells, Int. J. Oncol., 49 (2016), 529–538. https://doi.org/10.3892/ijo.2016.3555 doi: 10.3892/ijo.2016.3555
|
| [15] |
G. Cali, L. Insabato, D. Conza, G. Bifulco, L. Parrillo, P. Mirra, et al., GRP78 mediates cell growth and invasiveness in endometrial cancer, J. Cell. Physiol., 229 (2014), 1417–1426. https://doi.org/10.1002/jcp.24578 doi: 10.1002/jcp.24578
|
| [16] |
K. Matsuo, M. J. Gray, D. Y. Yang, S. A. Srivastava, P. B. Tripathi, L. A. Sonoda, et al., The endoplasmic reticulum stress marker, glucose-regulated protein-78 (GRP78) in visceral adipocytes predicts endometrial cancer progression and patient survival, Gynecol. Oncol., 128 (2013), 552–559. https://doi.org/10.1016/j.ygyno.2012.11.024 doi: 10.1016/j.ygyno.2012.11.024
|
| [17] |
D. Hui, C. E. Paiva, F. E. G. Del, C. Steer, J. Naberhuis, W. M. van de, et al., Prognostication in advanced cancer: update and directions for future research, Support Care Cancer, 27 (2019), 1973–1984. https://doi.org/10.1007/s00520-019-04727-y doi: 10.1007/s00520-019-04727-y
|
| [18] |
D. Hui, J. P. Maxwell, C. E. Paiva, Dealing with prognostic uncertainty: the role of prognostic models and websites for patients with advanced cancer, Curr. Opin. Support Palliat. Care, 13 (2019), 360–368. https://doi.org/10.1097/SPC.0000000000000459 doi: 10.1097/SPC.0000000000000459
|
| [19] |
M. E. Ritchie, B. Phipson, D. Wu, Y. Hu, C. W. Law, W. Shi, et al., limma powers differential expression analyses for RNA-sequencing and microarray studies, Nucleic Acids Res., 43 (2015), e47. https://doi.org/10.1093/nar/gkv007 doi: 10.1093/nar/gkv007
|
| [20] |
W. Walter, F. Sánchez-Cabo, M. Ricote, GOplot: an R package for visually combining expression data with functional analysis, Bioinformatics, 31 (2015), 2912–2914. https://doi.org/10.1093/bioinformatics/btv300 doi: 10.1093/bioinformatics/btv300
|
| [21] |
K. Ito, D. Murphy, Application of ggplot2 to pharmacometric graphics, CPT: Pharmacomet. Syst. Pharmacol., 2 (2013), e79. https://doi.org/10.1038/psp.2013.56 doi: 10.1038/psp.2013.56
|
| [22] |
G. Yu, L. G. Wang, Y. Han, Q. Y. He, ClusterProfiler: An R package for comparing biological themes among gene clusters, OMICS, 16 (2012), 284–287. https://doi.org/10.1089/omi.2011.0118 doi: 10.1089/omi.2011.0118
|
| [23] |
R. Tibshirani, The lasso method for variable selection in the Cox model, Stat. Med., 16 (1997), 385–395. https://doi.org/10.1002/(sici)1097-0258(19970228)16:4<385::aid-sim380>3.0.co;2-3 doi: 10.1002/(sici)1097-0258(19970228)16:4<385::aid-sim380>3.0.co;2-3
|
| [24] |
L. Li, Z. P. Liu, Detecting prognostic biomarkers of breast cancer by regularized Cox proportional hazards models, J. Transl. Med., 19 (2021), 514. https://doi.org/10.1186/s12967-021-03180-y doi: 10.1186/s12967-021-03180-y
|
| [25] |
P. Blanche, J. F. Dartigues, H. Jacqmin-Gadda, Estimating and comparing time-dependent areas under receiver operating characteristic curves for censored event times with competing risks, Stat. Med., 32 (2013), 5381–5397. https://doi.org/10.1002/sim.5958 doi: 10.1002/sim.5958
|
| [26] |
Y. Yang, H. J. Sun, Y. Zhang, T. F. Zhang, J. L. Gong, Y. B. Wei, et al., Dimensionality reduction by UMAP reinforces sample heterogeneity analysis in bulk transcriptomic data, Cell Rep., 36 (2021), 109442. https://doi.org/10.1016/j.celrep.2021.109442 doi: 10.1016/j.celrep.2021.109442
|
| [27] |
V. P. Balachandran, M. G. Mithat, J. J. Smith, R. P. DeMatteo, Nomograms in oncology: more than meets the eye, Lancet Oncol., 16 (2015), e173–e180. https://doi.org/10.1016/s1470-2045(14)71116-7 doi: 10.1016/s1470-2045(14)71116-7
|
| [28] |
U. B. Mogensen, H. Ishwaran, A. G. Thomas, Evaluating rrandom forests for survival analysis using prediction error ccurves, J. Statist. Software, 50 (2012), 1–23. https://doi.org/10.18637/jss.v050.i11 doi: 10.18637/jss.v050.i11
|
| [29] | A. Subramanian, P. Tamayo, V. K. Mootha, S. Mukherjee, B. L. Ebert, M. A. Gillette, et al., Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. U. S. A., 102 (2005), 15545–15550. https://doi.org/10.1073/pnas.0506580102 |
| [30] |
H. Wickham, The split-apply-combine srategy for data analysis, J. Statist. Software, 40 (2011), 1–29. https://doi.org/10.18637/jss.v040.i01 doi: 10.18637/jss.v040.i01
|
| [31] |
K. Yoshihara, M. Shahmoradgoli, E. Martinez, R. Vegesna, H. Kim, W. Torres-Garcia, et al., Inferring tumour purity and stromal and immune cell admixture from expression data, Nat. Commun., 4 (2013), 2612. https://doi.org/10.1038/ncomms3612 doi: 10.1038/ncomms3612
|
| [32] |
A. M. Newman, C. B. Steen, C. L. Liu, A. J. Gentles, A. A. Chaudhuri, F. Scherer, et al., Determining cell type abundance and expression from bulk tissues with digital cytometry, Nat. Biotechnol., 37 (2019), 773–782. https://doi.org/10.1038/s41587-019-0114-2 doi: 10.1038/s41587-019-0114-2
|
| [33] | S.Hänzelmann, R. Castelo, J. Guinney, GSVA: gene set variation analysis for microarray and RNA-seq data, BMC Bioinf., 14 (2013). https://doi.org/10.1186/1471-2105-14-7 |
| [34] |
P. Geeleher, N. Cox, R. S. Huang, pRRophetic: an R package for prediction of clinical chemotherapeutic response from tumor gene expression levels, PLoS One, 9 (2014), e107468. https://doi.org/10.1371/journal.pone.0107468 doi: 10.1371/journal.pone.0107468
|
| [35] |
J. Lamb, E. D. Crawford, D. Peck, J. W. Model, I. C. Bla, M. J. Wrobel, et al., The connectivity map: using gene-expression signatures to connect small molecules, genes, and disease, Science, 313 (2006), 1929–1935. https://doi.org/10.1126/science.1132939 doi: 10.1126/science.1132939
|
| [36] |
C. R. Smittenaar, K. A. Petersen, K. Stewart, N. Moitt, Cancer incidence and mortality projections in the UK until 2035, Br. J. Cancer, 115 (2016), 1147–1155. https://doi.org/10.1038/bjc.2016.304 doi: 10.1038/bjc.2016.304
|
| [37] |
J. Y. Park, J. Y. Lee, Y. Y. Lee, S. H. Shim, D. H. Suh, J. W. Kim, Major clinical research advances in gynecologic cancer in 2021, J. Gynecol. Oncol., 33 (2022), e43. https://doi.org/10.3802/jgo.2022.33.e43 doi: 10.3802/jgo.2022.33.e43
|
| [38] |
J. S. Ferriss, B. K. Erickson, I. M. Shih, A. N. Fader, Uterine serous carcinoma: key advances and novel treatment approaches, Int. J. Gynecol. Cancer, 31 (2021), 1165–1174. https://doi.org/10.1136/ijgc-2021-002753 doi: 10.1136/ijgc-2021-002753
|
| [39] |
S. A. Oakes, Endoplasmic reticulum stress signaling in cancer cells, Am. J. Pathol., 190 (2020), 934–946. https://doi.org/10.1016/j.ajpath.2020.01.010 doi: 10.1016/j.ajpath.2020.01.010
|
| [40] |
S. K. Hsu, C. C. Chiu, H. U. Dahms, C. K. Chou, C. M. Cheng, W. T. Chang, et al., Unfolded Protein Response (UPR) in survival, dormancy, immunosuppression, metastasis, and treatments of cancer cells, Int. J. Mol. Sci., 20 (2019), 2518. https://doi.org/10.3390/ijms20102518 doi: 10.3390/ijms20102518
|
| [41] |
D. Liang, M. Khoonkari, T. Avril, E. Chevet, F. A. E. Kruyt, The unfolded protein response as regulator of cancer stemness and differentiation: Mechanisms and implications for cancer therapy, Biochem. Pharmacol., 192 (2021), 114737. https://doi.org/10.1016/j.bcp.2021.114737 doi: 10.1016/j.bcp.2021.114737
|
| [42] |
X. Xu, Y. Q. Tang, J. S. Zhu, J. H. Luo, Endoplasmic reticulum stress-related four-biomarker risk classifier for survival evaluation in esophageal cancer, J. Oncol., 2022 (2022), 5860671. https://doi.org/10.1155/2022/5860671 doi: 10.1155/2022/5860671
|
| [43] |
R. Y. Huang, G. Z. Li, K. Y. Wang, Z. L. Wang, F. Zeng, H. M. Hu, et al., Comprehensive analysis of the clinical and biological significances of endoplasmic reticulum stress in diffuse gliomas, Front. Cell Dev. Biol., 9 (2021), 619396. https://doi.org/10.3389/fcell.2021.619396 doi: 10.3389/fcell.2021.619396
|
| [44] |
Q. Zhang, G. F. Guan, P. Cheng, W. Cheng, L. H. Yang, A. H. Wu, Characterization of an endoplasmic reticulum stress-related signature to evaluate immune features and predict prognosis in glioma, J. Cell. Mol. Med., 25 (2021), 3870–3884. https://doi.org/10.1111/jcmm.16321 doi: 10.1111/jcmm.16321
|
| [45] |
P. Liu, J. H. Wei, F. Y. Mao, Z. C. Xin, H. Duan, Y. Du, et al., Establishment of a prognostic model for hepatocellular carcinoma based on endoplasmic reticulum stress-related gene analysis, Front. Oncol., 11 (2021), 641487. https://doi.org/10.3389/fonc.2021.641487 doi: 10.3389/fonc.2021.641487
|
| [46] |
L. M. Morton, S. S. Wang, A. W. Bergen, N. Chatterjee, P. Kvale, R. Welch, et al., DRD2 genetic variation in relation to smoking and obesity in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial, Pharmacogenet. Genom., 16 (2006), 901–910. https://doi.org/10.1097/01.fpc.0000230417.20468.d0 doi: 10.1097/01.fpc.0000230417.20468.d0
|
| [47] |
F. Gemignani, S. Landi, V. Moreno, L. Gioia-Patricola, A. Chabrier, E. Guino, et al., Polymorphisms of the dopamine receptor gene DRD2 and colorectal cancer risk, Cancer Epidemiol Biomarkers Prev., 14 (2005), 1633–1638. https://doi.org/10.1158/1055-9965.EPI-05-0057 doi: 10.1158/1055-9965.EPI-05-0057
|
| [48] |
Y. Q. Tan, R. Sun, L. Liu, D. J. Yang, Q. Xiang, L. Li, et al., Tumor suppressor DRD2 facilitates M1 macrophages and restricts NF-kappaB signaling to trigger pyroptosis in breast cancer, Theranostics, 11 (2021), 5214–5231. https://doi.org/10.7150/thno.58322 doi: 10.7150/thno.58322
|
| [49] |
M. J. Mosquera, S. Kim, R. Bareja, Z. Fang, S. Y. Cai, H. Pan, et al., Extracellular matrix in synthetic hydrogel-based prostate cancer organoids regulate therapeutic response to EZH2 and DRD2 inhibitors, Adv. Mater., 34 (2022), e2100096. https://doi.org/10.1002/adma.202100096 doi: 10.1002/adma.202100096
|
| [50] |
L. He, K. Bhat, A. Ioannidis, L. Zhang, N. T. Nguyen, J. E. Allen, et al., Effects of the DRD2/3 antagonist ONC201 and radiation in glioblastoma, Radiother. Oncol., 161 (2021), 140–147. https://doi.org/10.1016/j.radonc.2021.05.027 doi: 10.1016/j.radonc.2021.05.027
|
| [51] |
J. Z. Liu, Y. X. Wei, Y. S. Wu, J. Li, J. Z. Sun, G. S. Ren, et al., ATP2C2 has potential to define tumor microenvironment in breast cancer, Front. Immunol., 12 (2021), 657950. https://doi.org/10.3389/fimmu.2021.657950 doi: 10.3389/fimmu.2021.657950
|
| [52] |
X. Chen, H. Y. Xie, X. Wang, Z. N. Zheng, S. Q. Jin, CIRBP knockdown attenuates tumourigenesis and improves the chemosensitivity of pancreatic cancer via the downregulation of DYRK1B, Front. Cell Dev. Biol., 9 (2021), 667551. https://doi.org/10.3389/fcell.2021.667551 doi: 10.3389/fcell.2021.667551
|
| [53] |
S. T. Boyle, V. Poltavets, J. Kular, N. T. Pyne, J. J. Sandow, A. C. Lewis, et al., ROCK-mediated selective activation of PERK signalling causes fibroblast reprogramming and tumour progression through a CRELD2-dependent mechanism, Nat. Cell. Biol., 22 (2020), 882–895. https://doi.org/10.1038/s41556-020-0523-y doi: 10.1038/s41556-020-0523-y
|
| [54] |
J. J. A. Marin, M. S. Garcia, G. X. Matias, J. M. Piulats, Facts and hopes in immunotherapy of endometrial cancer, Clin. Cancer Res., 28 (2022), 4849–4860. https://doi.org/10.1158/1078-0432.CCR-21-1564 doi: 10.1158/1078-0432.CCR-21-1564
|
| [55] |
R. Voelker, Immunotherapy approved for endometrial cancer, JAMA, 325 (2021), 2143. https://doi.org/10.1001/jama.2021.8319 doi: 10.1001/jama.2021.8319
|
| [56] |
J. H. Liu, Y. C. Wang, J. Mei, S. P. Nie, Y. Zhang, Identification of a novel immune landscape signature for predicting prognosis and response of endometrial carcinoma to immunotherapy and chemotherapy, Front. Cell Dev. Biol., 9 (2021), 671736. https://doi.org/10.3389/fcell.2021.671736 doi: 10.3389/fcell.2021.671736
|
| [57] |
G. W. Wang, D. D. Wang, M. G. Sun, X. F. Liu, Q. Yang, Identification of prognostic and immune-related gene signatures in the tumor microenvironment of endometrial cancer, Int. Immunopharmacol., 88 (2020), 106931. https://doi.org/10.1016/j.intimp.2020.106931 doi: 10.1016/j.intimp.2020.106931
|
| [58] | Y. Meng, Y. B. Yang, Y. Zhang, X. H. Yang, X. M. Li, C. Hu, The role of an immune signature for prognosis and immunotherapy response in endometrial cancer, Am. J. Transl. Res., 13 (2021), 532–548. |
| [59] |
J. Hu, J. Sun, MUC16 mutations improve patients' prognosis by enhancing the infiltration and antitumor immunity of cytotoxic T lymphocytes in the endometrial cancer microenvironment, Oncoimmunology, 7 (2018), e1487914. https://doi.org/10.1080/2162402X.2018.1487914 doi: 10.1080/2162402X.2018.1487914
|
| [60] |
T. Kolben, M. Mannewitz, C. Perleberg, K. Schnell, D. Anz, L. Hahn, et al., Presence of regulatory T-cells in endometrial cancer predicts poorer overall survival and promotes progression of tumor cells, Cell Oncol. (Dordr), 2022. https://doi.org/10.1007/s13402-022-00708-2 doi: 10.1007/s13402-022-00708-2
|
| [61] | W. Y. Cao, X. Y. Ma, J. V. Fischer, C. G. Sun, B. H. Kong, Q. Zhang, Immunotherapy in endometrial cancer: rationale, practice and perspectives, Biomark. Res., 9 (2021), 49. https://doi.org/10.1186/s40364-021-00301-z |
| [62] |
E. Wang, A. Sorolla, Sensitizing endometrial cancer to ionizing radiation by multi-tyrosine kinase inhibition, J. Gynecol. Oncol., 31 (2020), e29. https://doi.org/10.3802/jgo.2020.31.e29 doi: 10.3802/jgo.2020.31.e29
|
| [63] |
V. Makker, N. Colombo, H. A. Casado, A. D. Santin, E. Colomba, D. S. Miller, et al., Lenvatinib plus pembrolizumab for advanced endometrial cancer, N. Engl. J. Med., 386 (2022), 437–448. https://doi.org/10.1056/NEJMoa2108330 doi: 10.1056/NEJMoa2108330
|
 mbe-20-02-188-Suppplementary Figures 1 and 2.pdf mbe-20-02-188-Suppplementary Figures 1 and 2.pdf |
 |
Figures(7) / Tables(3)

Pei Zhou, Caiyun Wu, Cong Ma, Ting Luo, Jing Yuan, Ping Zhou, Zhaolian Wei. Identification of an endoplasmic reticulum stress-related gene signature to predict prognosis and potential drugs of uterine corpus endometrial cancer[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 4018-4039. doi: 10.3934/mbe.2023188







 DownLoad:
DownLoad: