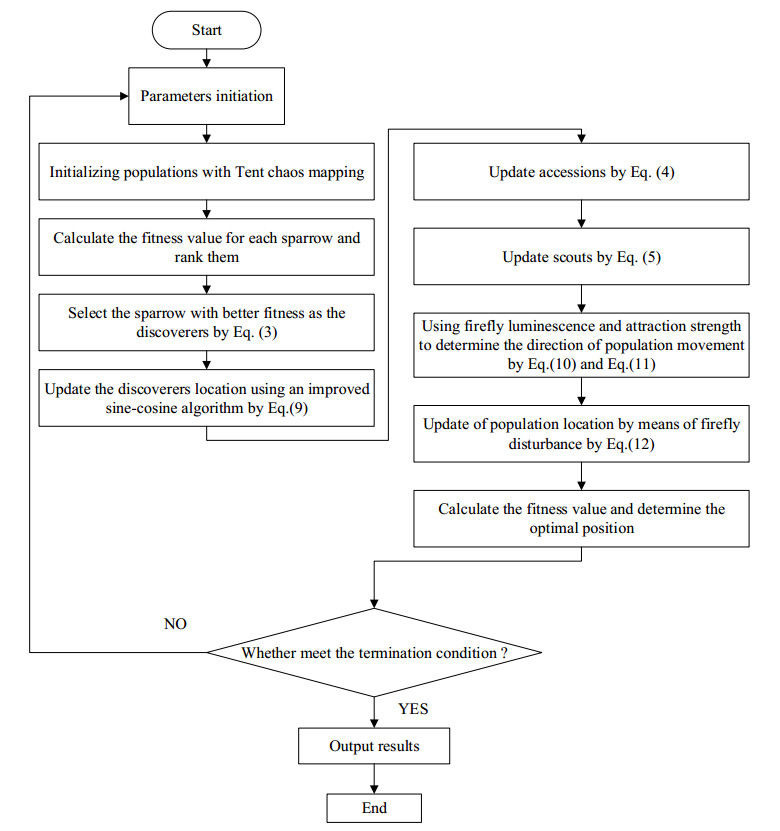
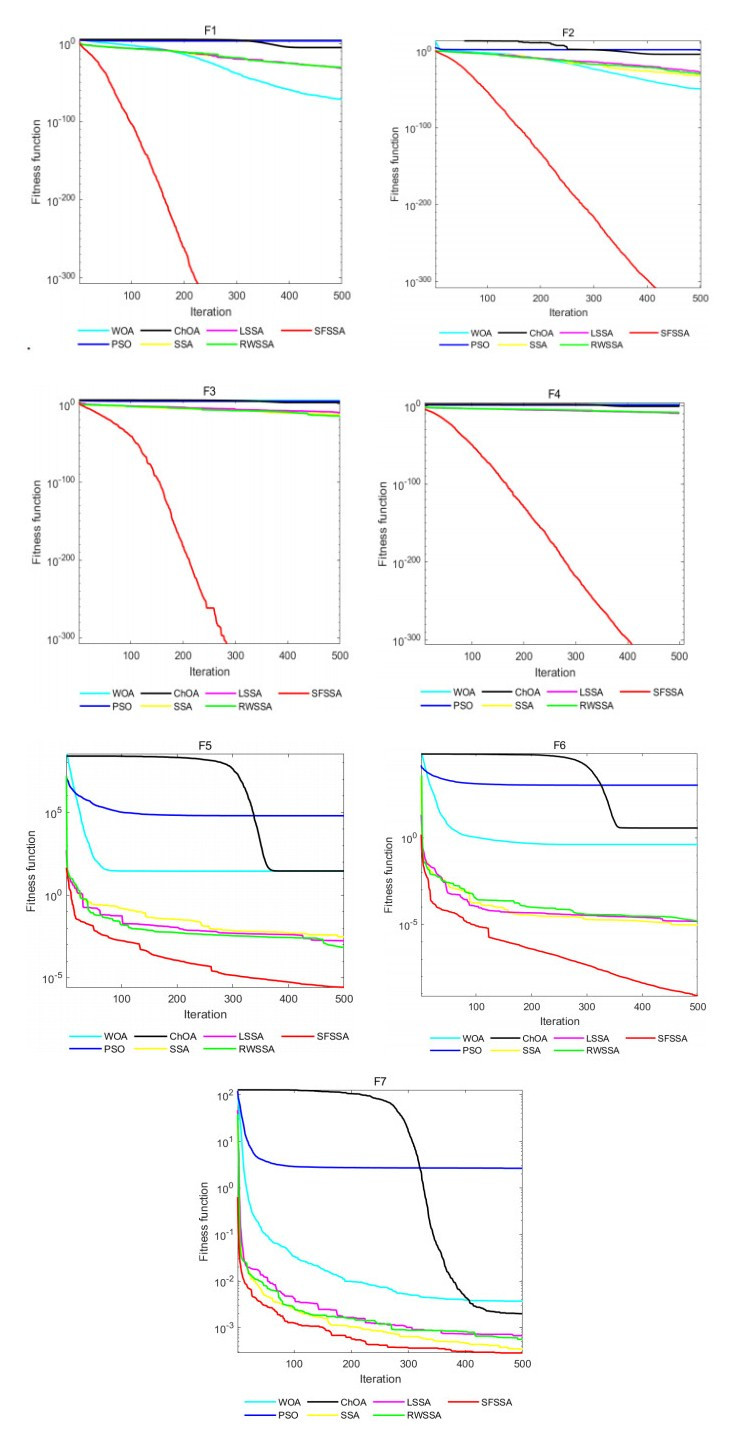
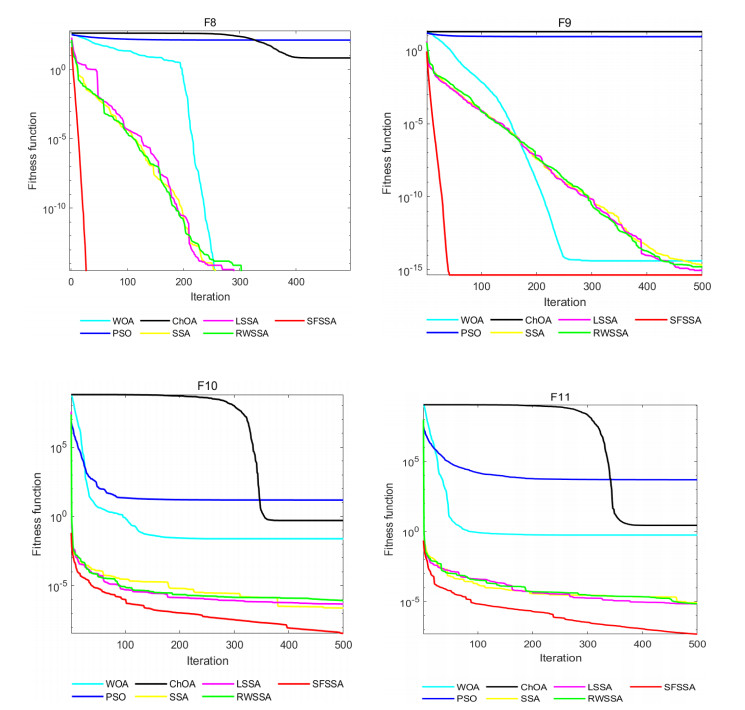
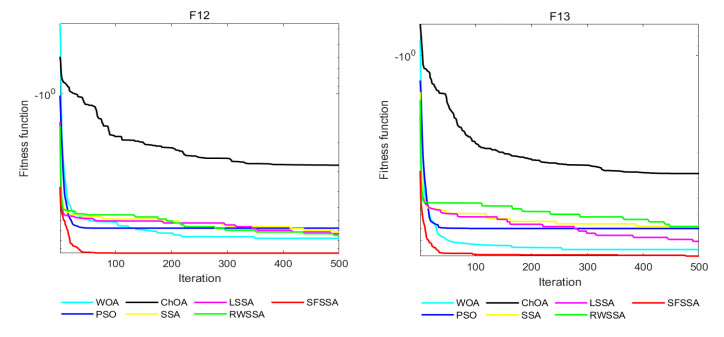
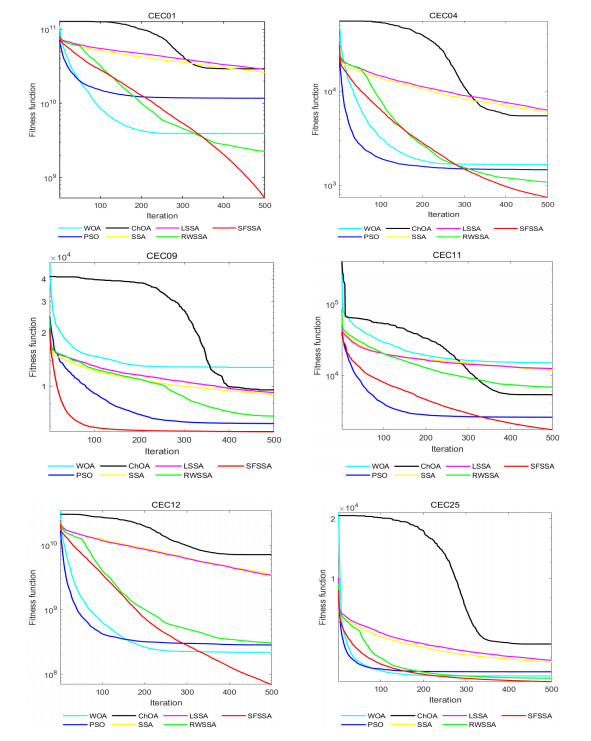
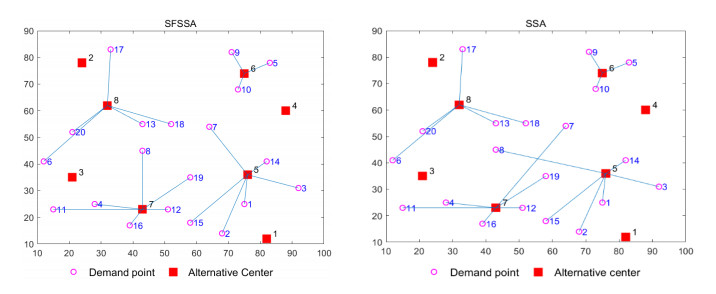
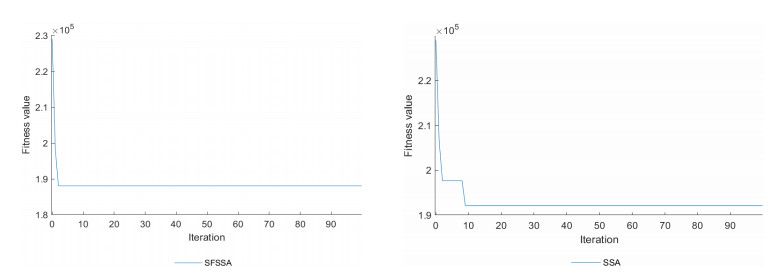
Swarm intelligence algorithms are relatively simple and highly applicable algorithms, especially for solving optimization problems with high reentrancy, high stochasticity, large scale, multi-objective and multi-constraint characteristics. The sparrow search algorithm (SSA) is a kind of swarm intelligence algorithm with strong search capability, but SSA has the drawback of easily falling into local optimum in the iterative process. Therefore, a sine cosine and firefly perturbed sparrow search algorithm (SFSSA) is proposed for addressing this deficiency. Firstly, the Tent chaos mapping is invoked in the initialization population stage to improve the population diversity; secondly, the positive cosine algorithm incorporating random inertia weights is introduced in the discoverer position update, so as to improve the probability of the algorithm jumping out of the local optimum and speed up the convergence; finally, the firefly perturbation is used to firefly perturb the sparrows, and all sparrows are updated with the optimal sparrows using the firefly perturbation method to improve their search-ability. Thirteen benchmark test functions were chosen to evaluate SFSSA, and the results were compared to those computed by existing swarm intelligence algorithms, as well as the proposed method was submitted to the Wilcoxon rank sum test. Furthermore, the aforesaid methods were evaluated in the CEC 2017 test functions to further validate the optimization efficiency of the algorithm when the optimal solution is not zero. The findings show that SFSSA is more favorable in terms of algorithm performance, and the method's searchability is boosted. Finally, the suggested algorithm is used to the locating problem of emergency material distribution centers to further validate the feasibility and efficacy of SFSSA.
Citation: Xiangyang Ren, Shuai Chen, Kunyuan Wang, Juan Tan. Design and application of improved sparrow search algorithm based on sine cosine and firefly perturbation[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 11422-11452. doi: 10.3934/mbe.2022533
Swarm intelligence algorithms are relatively simple and highly applicable algorithms, especially for solving optimization problems with high reentrancy, high stochasticity, large scale, multi-objective and multi-constraint characteristics. The sparrow search algorithm (SSA) is a kind of swarm intelligence algorithm with strong search capability, but SSA has the drawback of easily falling into local optimum in the iterative process. Therefore, a sine cosine and firefly perturbed sparrow search algorithm (SFSSA) is proposed for addressing this deficiency. Firstly, the Tent chaos mapping is invoked in the initialization population stage to improve the population diversity; secondly, the positive cosine algorithm incorporating random inertia weights is introduced in the discoverer position update, so as to improve the probability of the algorithm jumping out of the local optimum and speed up the convergence; finally, the firefly perturbation is used to firefly perturb the sparrows, and all sparrows are updated with the optimal sparrows using the firefly perturbation method to improve their search-ability. Thirteen benchmark test functions were chosen to evaluate SFSSA, and the results were compared to those computed by existing swarm intelligence algorithms, as well as the proposed method was submitted to the Wilcoxon rank sum test. Furthermore, the aforesaid methods were evaluated in the CEC 2017 test functions to further validate the optimization efficiency of the algorithm when the optimal solution is not zero. The findings show that SFSSA is more favorable in terms of algorithm performance, and the method's searchability is boosted. Finally, the suggested algorithm is used to the locating problem of emergency material distribution centers to further validate the feasibility and efficacy of SFSSA.

| [1] |
L. Brezočnik, I. Fister, V. Podgorelec, Swarm intelligence algorithms for feature selection: a review, Appl. Sci., 8 (2018), 1521. https://doi.org/10.3390/app8091521 doi: 10.3390/app8091521

|
| [2] |
Y. L. Li, S. Q. Wang, Q. R. Chen, X. G. Wang, Comparative study of several new swarm intelligence optimization algorithms, Comput. Eng. Appl., 56 (2020), 1–12. https://doi.org/10.3778/j.issn.1002-8331.2006-0291 doi: 10.3778/j.issn.1002-8331.2006-0291
|
| [3] | J. Kennedy, R. Eberhart, Particle swarm optimization, in Proceedings of the ICNN'95-international conference on neural networks, 4 (1995), 1942–1948. https://doi.org/10.1109/ICNN.1995.488968 |
| [4] | R. Eberhart, J. Kennedy, A new optimizer using particle swarm theory, in Proceedings of the MHS'95 6th International Symposium on Micro Machine and Human Science, IEEE, (1995), 39–43. https://doi.org/10.1109/MHS.1995.494215 |
| [5] |
M. Dorigo, V. Maniezzo, A. Colorni, Ant system: optimization by a colony of cooperating agents, IEEE Trans. Syst. Man Cybern. Part B Cybern., 26 (1996), 29–41. https://doi.org/10.1109/3477.484436 doi: 10.1109/3477.484436
|
| [6] |
M. Dorigo, L. M. Gambardella, Ant colony system: a cooperative learning approach to the traveling salesman problem, IEEE Trans. Evol. Comput., 1 (1997), 53–66. https://doi.org/10.1109/4235.585892 doi: 10.1109/4235.585892
|
| [7] | X. L. Li, A New Intelligent Optimization Method-Artificial Fish Swarm Algorithm, Ph.D thesis, Zhejiang University, Hangzhou, 2003. https://doi.org/cnki.cdmd.1.2003.051212 |
| [8] |
S. Mirjalili, S. M. Mirjalili, A. Lewis, Grey wolf optimizer, Adv. Eng. Software, 69 (2014), 46–61. https://doi.org/10.1016/j.advengsoft.2013.12.007 doi: 10.1016/j.advengsoft.2013.12.007
|
| [9] |
S. Mirjalili, A. Lewis, The whale optimization algorithm, Adv. Eng. Software, 95 (2016), 51–67. https://doi.org/10.1016/j.advengsoft.2016.01.008 doi: 10.1016/j.advengsoft.2016.01.008
|
| [10] | X. S. Yang, A new metaheuristic bat-inspired algorithm, Comput. Knowl. Technol., 284 (2010), 65–74. https://doi.org/arXiv-1004.4170 |
| [11] |
I. Fister, I. Fister Jr., X. S. Yang, J. Brest, A comprehensive review of firefly algorithms, Swarm Evol. Comput., 13 (2013), 34–46. https://doi.org/10.1016/j.swevo.2013.06.001 doi: 10.1016/j.swevo.2013.06.001
|
| [12] |
M. Khishe, M. R. Mosavi, Chimp optimization algorithm, Expert systems with applications, 149 (2020), 113338. https://doi.org/10.1016/j.eswa.2020.113338 doi: 10.1016/j.eswa.2020.113338
|
| [13] |
J. K. Xue, B. Shen, A novel swarm intelligence optimization approach: sparrow search algorithm, Syst. Sci. Control Eng., 8 (2020), 22–34. https://doi.org/10.1080/21642583.2019.1708830 doi: 10.1080/21642583.2019.1708830
|
| [14] |
Q. C. Duan, M. X. Mao, P. Duan, B. Hu, An improved artificial fish swarm algorithm optimized by particle swarm optimization algorithm with extended memory, Kybernetes, 45 (2016). https://doi.org/10.1108/k-09-2014-0198 doi: 10.1108/k-09-2014-0198
|
| [15] |
Z. L. Han, M. Chen, S. Y. Shao, Q. X. Wu, Improved artificial bee colony algorithm-based path planning of unmanned autonomous helicopter using multi-strategy evolutionary learning, Aerosp. Sci. Technol., 122 (2022), 107374. https://doi.org/10.1016/j.ast.2022.107374 doi: 10.1016/j.ast.2022.107374
|
| [16] |
S. Dereli, A novel approach based on average swarm intelligence to improve the whale optimization algorithm, Arabian J. Sci. Eng., 47 (2022), 1763–1776. https://doi.org/10.1007/s13369-021-06042-3 doi: 10.1007/s13369-021-06042-3
|
| [17] |
R. Chaudhary, H. Banati, Swarm bat algorithm with improved search (SBAIS), Soft Comput., 23 (2019), 11461–11491. https://doi.org/10.1007/s00500-018-03688-4 doi: 10.1007/s00500-018-03688-4
|
| [18] |
J. R. Wu, Y. G. Wang, K. Burrang, Y. C. Tian, B. Lawson, Z. Ding, An improved firefly algorithm for global continuous optimization problems, Expert Syst. Appl., 149 (2020), 113340. https://doi.org/10.1016/j.eswa.2020.113340 doi: 10.1016/j.eswa.2020.113340
|
| [19] |
X. Lv, X. D. Mu, J. Zhang, Z. Wang, Chaos sparrow search optimization algorithm, J. Beijing Univ. Aeronaut. Astronaut., 47 (2020), 1–10. https://doi.org/10.13700/j.bh.1001-5965.2020.0298 doi: 10.13700/j.bh.1001-5965.2020.0298
|
| [20] |
B. Ma, P. M. Lu, L. F. Zhang, Y. Liu, Q. Zhou, Y. Chen, et al., Enhanced sparrow search algorithm with mutation strategy for global optimization, IEEE Access, 9 (2021), 159218–159261. https://doi.org/10.1109/access.2021.3129255 doi: 10.1109/access.2021.3129255
|
| [21] |
J. Ma, Z. Y. Hao, W. J. Sun, Enhancing sparrow search algorithm via multi-strategies for continuous optimization problems, Inf. Process. Manage., 59 (2022), 102854. https://doi.org/10.1016/j.ipm.2021.102854 doi: 10.1016/j.ipm.2021.102854
|
| [22] |
C. Y. Wu, X. S. Fu, J. K. Pei, Z. G. Dong, A novel sparrow search algorithm for the traveling salesman problem, IEEE Access, 9 (2021), 153456–153471. https://doi.org/10.1109/access.2021.3128433 doi: 10.1109/access.2021.3128433
|
| [23] |
Z. Zhang, R. He, K. Yang, A bioinspired path planning approach for mobile robots based on improved sparrow search algorithm, Adv. Manuf., 10 (2022), 114–130. https://doi.org/10.1007/s40436-021-00366-x doi: 10.1007/s40436-021-00366-x
|
| [24] |
Y. Yu, S. C. Gao, S. Chen, S. Cheng, Y. R. Wang, S. Y. Song, et al., CBSO: a memetic brain storm optimization with chaotic local search, Memetic Comput., 10 (2018), 353–367. https://doi.org/10.1007/s12293-017-0247-0 doi: 10.1007/s12293-017-0247-0
|
| [25] |
E. V. Altay, B. Alatas, Bird swarm algorithms with chaotic mapping, Artif. Intell. Rev., 53 (2020), 1373–1414. https://doi.org/10.1007/s10462-019-09704-9 doi: 10.1007/s10462-019-09704-9
|
| [26] |
L. F. Yue, R. N. Yang, Y. J. Zhang, Y. Yu, Z. X. Zhang, Tent chaos and simulated annealing improved moth-flame optimization algorithm, J. Harbin Inst. Technol., 51 (2019), 146–154. https://doi.org/10.11918/j.issn.0367-6234.201811027 doi: 10.11918/j.issn.0367-6234.201811027
|
| [27] |
S. R. Chen, S. Y. Wang, An optimization method for an integrated energy system scheduling process based on NSGA-Ⅱ improved by tent mapping chaotic algorithms, Processes, 8 (2020), 426. https://doi.org/10.3390/pr8040426 doi: 10.3390/pr8040426
|
| [28] |
F. B. Demir, T. Tuncer, A. F. Kocamaz, A chaotic optimization method based on logistic-sine map for numerical function optimization, Neural Comput. Appl., 32 (2020), 14227–14239. https://doi.org/10.1007/s00521-020-04815-9 doi: 10.1007/s00521-020-04815-9
|
| [29] |
Z. J. Teng, J. L. Lv, L. W. Guo, Y. Y. Xu, An improved hybrid grey wolf optimization algorithm based on Tent mapping, J. Harbin Inst. Technol., 50 (2018), 40–49. https://doi.org/10.11918/j.issn.0367-6234.201806096 doi: 10.11918/j.issn.0367-6234.201806096
|
| [30] |
X. F. Zhang, F. Xiao, X. L. Tong, J. Yun, Y. Liu, Y. Sun, et al., Time optimal trajectory planing based on improved Sparrow Search Algorithm, Front. Bioeng. Biotechnol. , 10 (2022), 852408. https://doi.org/10.3389/fbioe.2022.852408 doi: 10.3389/fbioe.2022.852408
|
| [31] |
J. M. Liu, W, Zhang, S. S. Qiu, Method for improving randomness of Tent chaotic sequences, Comput. Eng., 38 (2012), 10–12. https://doi.org/10.3969/j.issn.1000-3428.2012.06.003 doi: 10.3969/j.issn.1000-3428.2012.06.003
|
| [32] |
D. Zhao, Y. He, Chaotic binary bat algorithm for analog test point selection, Analog Integr. Circuits Signal Process., 84 (2015), 201–214. https://doi.org/10.1007/s10470-015-0548-5 doi: 10.1007/s10470-015-0548-5
|
| [33] |
S. Mirjalili, SCA: A Sine Cosine Algorithm for solving optimization problems, Knowl.-Based Syst., 96 (2016), 120–133. https://doi.org/10.1016/j.knosys.2015.12.022 doi: 10.1016/j.knosys.2015.12.022
|
| [34] |
C. Gan, W. H. Cao, M. Wu, X. Chen, A new bat algorithm based on iterative local search and stochastic inertia weight, Expert Syst. Appl., 104 (2018), 202–212. https://doi.org/10.1016/j.eswa.2018.03.015 doi: 10.1016/j.eswa.2018.03.015
|
| [35] |
A. H. Gandomi, X. S. Yang, S. Talatahari, A. H. Alavi, Firefly algorithm with chaos, Commun. Nonlinear Sci. Numer. Simul., 18 (2013), 89–98. https://doi.org/10.1016/j.cnsns.2012.06.009 doi: 10.1016/j.cnsns.2012.06.009
|
| [36] |
D. L. Zhang, H. W. Xia, C. X. Chao, G. C. Ma, C. H. Wang, Improved firefly algorithm and its convergence analysis, Syst. Eng. Electron., 44 (2022), 1291–1300. https://doi.org/10.12305/j.issn.1001-506X.2022.04.27 doi: 10.12305/j.issn.1001-506X.2022.04.27
|
| [37] |
A. Yelghi, C. Köse, A modified firefly algorithm for global minimum optimization, Appl. Soft Comput., 62 (2018), 29–44. https://doi.org/10.1016/j.asoc.2017.10.032 doi: 10.1016/j.asoc.2017.10.032
|
| [38] |
W. Long, T. B. Wu, S. H. Cai, X. M. Liang, J. J. Jiao, M. Xu, A novel grey wolf optimizer algorithm with refraction learning, IEEE Access, 7 (2019), 57805–57819. https://doi.org/10.1109/access.2019.2910813 doi: 10.1109/access.2019.2910813
|
| [39] |
J. Derrac, S. García, D. Molina, F. Herrera, A practical tutorial on the use of nonparametric statistical tests as a methodology for comparing evolutionary and swarm intelligence algorithms, Swarm Evol. Comput., 1 (2011), 3–18. https://doi.org/10.1016/j.swevo.2011.02.002 doi: 10.1016/j.swevo.2011.02.002
|
Figures(9) / Tables(13)

Xiangyang Ren, Shuai Chen, Kunyuan Wang, Juan Tan. Design and application of improved sparrow search algorithm based on sine cosine and firefly perturbation[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 11422-11452. doi: 10.3934/mbe.2022533







 DownLoad:
DownLoad: