The human tear film is rapidly established after each blink, and is essential for clear vision and eye health. This paper reviews mathematical models and theories for the human tear film on the ocular surface, with an emphasis on localized flows where the tear film may fail. The models attempt to identify the important physical processes, and their parameters, governing the tear film in health and disease.
Citation: Richard J. Braun, Rayanne A. Luke, Tobin A. Driscoll, Carolyn G. Begley. Dynamics and mechanisms for tear breakup (TBU) on the ocular surface[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5146-5175. doi: 10.3934/mbe.2021262
The human tear film is rapidly established after each blink, and is essential for clear vision and eye health. This paper reviews mathematical models and theories for the human tear film on the ocular surface, with an emphasis on localized flows where the tear film may fail. The models attempt to identify the important physical processes, and their parameters, governing the tear film in health and disease.

| [1] |
A. J. Bron, J. M. Tiffany, S. M. Gouveia, N. Yokoi, L. W. Voon, Functional aspects of the tear film lipid layer, Exp. Eye Res., 78 (2004), 347-360. doi: 10.1016/j.exer.2003.09.019

|
| [2] |
B. Govindarajan, I. K. Gipson, Membrane-tethered mucins have multiple functions on the ocular surface, Exp. Eye Res., 90 (2010), 655-693. doi: 10.1016/j.exer.2010.02.014
|
| [3] |
R. Montés-Micó, A. Cervino, T. Ferrer-Blasco, S. García-Lázaro, D. Madrid-Costa, The tear film and the optical quality of the eye, Ocul. Surf., 8 (2010), 185-192. doi: 10.1016/S1542-0124(12)70233-1
|
| [4] | R. Tutt, A. Bradley, C. G. Begley, L. N. Thibos, Optical and visual impact of tear break-up in human eyes, Invest. Ophthalmol. Vis. Sci., 41 (2000), 4117-4123. |
| [5] |
M. G. Doane, Blinking and the mechanics of the lacrimal drainage system, Ophthalmology, 88 (1981), 844-851. doi: 10.1016/S0161-6420(81)34940-9
|
| [6] |
S. Khanal, T. J. Millar, Nanoscale phase dynamics of the normal tear film, Nanomed.: Nanotech. Biol. Med., 6 (2010), 707-13. doi: 10.1016/j.nano.2010.06.002
|
| [7] |
S. Mishima, D. M. Maurice, The oily layer of the tear film and evaporation from the corneal surface, Exp. Eye Res., 1 (1961), 39-45. doi: 10.1016/S0014-4835(61)80006-7
|
| [8] | T. F. Svitova, M. C. Lin, Evaporation retardation by model tear-lipid films: The roles of film aging, compositions and interfacial rheological properties, Coll. Surf. B, 197 (2021), 111392. |
| [9] | M. S. Norn, Semiquantitative interference study of the fatty layer of precorneal film, Acta Ophthalmol., 57 (1979), 766-774. |
| [10] |
F. J. Holly, Formation and rupture of the tear film, Exp. Eye Res., 15 (1973), 515-525. doi: 10.1016/0014-4835(73)90064-X
|
| [11] | J. M. Tiffany, Measurement of wettability of the corneal epithelium I. particle attachment method, Acta Ophthalmol., 68 (1990), 175-181. |
| [12] | J. M. Tiffany, Measurement of wettability of the corneal epithelium II. contact angle method, Acta Ophthalmol., 68 (1990), 182-187. |
| [13] |
I. K. Gipson, Distribution of mucins at the ocular surface, Exp. Eye Res., 78 (2004), 379-388. doi: 10.1016/S0014-4835(03)00204-5
|
| [14] |
P. E. King-Smith, B. A. Fink, R. M. Hill, K. W. Koelling, J. M. Tiffany, The thickness of the tear film, Curr. Eye. Res., 29 (2004), 357-368. doi: 10.1080/02713680490516099
|
| [15] |
N. Yokoi, Y. Takehisa, S. Kinoshita, Correlation of tear lipid layer interference patterns with the diagnosis and severity of dry eye, Am. J. Ophthalmol., 122 (1996), 818-824. doi: 10.1016/S0002-9394(14)70378-2
|
| [16] |
E. Goto, S. C. G. Tseng, Kinetic analysis of tear interference images in aqueous tear deficiency dry eye before and after punctal occlusion, Invest. Ophthalmol. Vis. Sci., 44 (2003), 1897-1905. doi: 10.1167/iovs.02-0818
|
| [17] |
P. E. King-Smith, J. J. Nichols, R. J. Braun, K. K. Nichols, High resolution microscopy of the lipid layer of the tear film, Ocul. Surf., 9 (2011), 197-211. doi: 10.1016/S1542-0124(11)70033-7
|
| [18] |
R. J. Braun, P. E. King-Smith, C. G. Begley, Longfei Li, N. R. Gewecke, Dynamics and function of the tear film in relation to the blink cycle, Prog. Retin. Eye Res., 45 (2015), 132-164. doi: 10.1016/j.preteyeres.2014.11.001
|
| [19] |
A. Tomlinson, M. G. Doane, A. McFadyen, Inputs and outputs of the lacrimal system: review of production and evaporative loss, Ocul. Surf., 7 (2009), 186-198. doi: 10.1016/S1542-0124(12)70186-6
|
| [20] |
S. H. Kimball, P. E. King-Smith, J. J. Nichols, Evidence for the major contribution of evaporation to tear film thinning between blinks, Invest. Opthalmol. Vis. Sci., 51 (2010), 6294-6297. doi: 10.1167/iovs.09-4772
|
| [21] | R. J. Braun, Dynamics of the tear film, Annu. Rev. Fluid Mech., 44 (2012), 267-297. |
| [22] |
C. F. Cerretani, C. J. Radke, Tear dynamics in healthy and dry eyes, Curr. Eye Res., 39 (2014), 580-595. doi: 10.3109/02713683.2013.859274
|
| [23] |
M. A. Lemp, C. Baudouin, J. Baum, M. Dogru, G. N. Foulks, S. Kinoshita, et al., The definition and classification of dry eye disease, Ocul. Surf., 5 (2007), 75-92. doi: 10.1016/S1542-0124(12)70081-2
|
| [24] |
J. A. Smith, J. Albeitz, C. G. Begley, B. Caffrey, K. K. Nichols, D. Schaumberg, et al., The
epidemiology of dry eye disease: Report of the Epidemiology Subcommittee of the International
Dry Eye WorkShop, Ocul. Surf., 5 (2007), 93-107. doi: 10.1016/S1542-0124(12)70082-4
|
| [25] |
F. Stapleton, M. Alves, V. Y. Bunya, I. Jalbert, K. Lekhanont, F. Malet, et al., TFOS DEWS-II Epidemiology Report, Ocul. Surf., 15 (2017), 334-365. doi: 10.1016/j.jtos.2017.05.003
|
| [26] |
C. Baudouin, P. Aragona, E. M. Messmer, A. Tomlinson, M. Calonge, K. G. Boboridis, et al., Role of hyperosmolarity in the pathogenesis and management of dry eye disease: Proceedings of the OCEAN group meeting, Ocul. Surf., 11 (2013), 246-258. doi: 10.1016/j.jtos.2013.07.003
|
| [27] |
L. Li, R. J. Braun, W. D. Henshaw, P. E. King-Smith, Computed tear film and osmolarity dynamics on an eye-shaped domain, Math. Med. Biol., 33 (2016), 123-157. doi: 10.1093/imammb/dqv013
|
| [28] |
M. A. Lemp, A. J. Bron, C. Baudoin, J. M. Benitez Del Castillo, D. Geffen, J. Tauber, et al., Tear osmolarity in the diagnosis and management of dry eye disease, Am. J. Ophthalmol., 151 (2011), 792-798. doi: 10.1016/j.ajo.2010.10.032
|
| [29] |
A. Tomlinson, S. Khanal, K. Ramesh, C. Diaper, A. McFadyen, Tear film osmolarity as a referent for dry eye diagnosis, Invest. Ophthalmol. Vis. Sci., 47 (2006), 4309-4315. doi: 10.1167/iovs.05-1504
|
| [30] |
P. Versura, V. Profazio, E. C. Campos, Performance of tear osmolarity compared to previous diagnostic tests for dry eye diseases, Curr. Eye Res., 35 (2010), 553-564. doi: 10.3109/02713683.2010.484557
|
| [31] | N. W. Tietz, Clinical Guide to Laboratory Tests, 3rd edition, W. B. Saunders, 1995. |
| [32] |
D. A. Dartt, M. D. P. Willcox, Complexity of the tear film: Importance in homeostasis and dysfunction during disease, Exp. Eye Res., 117 (2013), 1-3. doi: 10.1016/j.exer.2013.10.008
|
| [33] |
J. P. Gilbard, R. L. Farris, J. Santamaria, Osmolarity of tear microvolumes in keratoconjunctivitis sicca, Arch. Ophthalmol., 96 (1978), 677-681. doi: 10.1001/archopht.1978.03910050373015
|
| [34] |
B. D. Sullivan, D. Whitmer, K. K. Nichols, A. Tomlinson, G. N. Foulks, G. Geerling, et al., An objective approach to dry eye disease severity, Invest. Ophthalmol. Vis. Sci., 51 (2010), 6125-6130. doi: 10.1167/iovs.10-5390
|
| [35] |
H. Liu, C. G. Begley, M. Chen, A. Bradley, J. Bonanno, N. A. McNamara, et al., A link between tear instability and hyperosmolarity in dry eye, Invest. Ophthalmol. Vis. Sci., 50 (2009), 3671-3679. doi: 10.1167/iovs.08-2689
|
| [36] |
C. C. Peng, C. Cerretani, R. J. Braun, C. J. Radke, Evaporation-driven instability of the precorneal tear film, Adv. Coll. Interface Sci., 206 (2014), 250-264. doi: 10.1016/j.cis.2013.06.001
|
| [37] |
M. G. Doane, An instrument for in vivo tear film interferometry, Optom. Vis. Sci., 66 (1989), 383-388. doi: 10.1097/00006324-198906000-00008
|
| [38] |
P. E. King-Smith, B. A. Fink, J. J. Nichols, K. K. Nichols, R. J. Braun, G. B. McFadden, The contribution of lipid layer movement to tear film thinning and breakup, Invest. Ophthalmol. Visual Sci., 50 (2009), 2747-2756. doi: 10.1167/iovs.08-2459
|
| [39] |
J. Wang, D. Fonn, T. L. Simpson, L. Jones, Precorneal and pre- and postlens tear film thickness measured indirectly with optical coherence tomography, Invest. Ophthalmol. Vis. Sci., 44 (2003), 2524-2528. doi: 10.1167/iovs.02-0731
|
| [40] |
C. G. Begley, T. Simpson, H. Liu, E Salvo, Z. Wu, A. Bradley, P. Situ, Quantative analysis of tear film fluorescence and discomfort during tear film instability and thinning, Invest. Ophthalmol. Vis. Sci., 54 (2013), 2645-2653. doi: 10.1167/iovs.12-11299
|
| [41] | D. A. Benedetto, T. E. Clinch, P. R. Laibson, In vivo observations of tear dynamics using fluorophotometry, Arch. Ophthalmol., 102 (1986), 410-412. |
| [42] | M. S. Norn, Dessication of the precorneal film I. Corneal wetting time, Acta Ophthalmol., 47 (1969), 865-880. |
| [43] |
P. E. King-Smith, P. Ramamoorthy, R. J. Braun, J. J. Nichols, Tear film images and breakup analyzed using fluorescent quenching, Invest. Ophthalmol. Vis. Sci., 54 (2013), 6003--6011. doi: 10.1167/iovs.13-12628
|
| [44] |
W. R. S. Webber, D. P. Jones, Continuous fluorophotometric method measuring tear turnover rate in humans and analysis of factors affecting accuracy, Med. Biol. Eng. Comput., 24 (1986), 386-392. doi: 10.1007/BF02442693
|
| [45] |
J. J. Nichols, P. E. King-Smith, E. A. Hinel, M. Thangavelu, K. K. Nichols, The use of fluorescent quenching in studying the contribution of evaporation to tear thinning, Invest. Ophthalmol. Visual Sci., 53 (2012), 5426-5432. doi: 10.1167/iovs.12-10033
|
| [46] |
R. J. Braun, N. Gewecke, C. G. Begley, P. E. King-Smith, J. I. Siddique, A model for tear film thinning with osmolarity and fluorescein, Invest. Ophthalmol. Vis. Sci., 55 (2014), 1133-1142. doi: 10.1167/iovs.13-12773
|
| [47] | R. J. Braun, T. A. Driscoll, C. G. Begley, Mathematical models of the tear film, in Ocular Fluid Dynamics, Springer, Berlin, (2019), 387-432. |
| [48] |
R. E. Berger, S. Corrsin, A surface tension gradient mechanism for driving the pre-corneal tear film after a blink, J. Biomech., 7 (1974), 225-238. doi: 10.1016/0021-9290(74)90013-X
|
| [49] |
R. J. Braun, R. Usha, G. B. McFadden, T. A. Driscoll, L. P. Cook, P. E. King-Smith, Thin film dynamics on a prolate spheroid with application to the cornea, J. Eng. Math., 73 (2012), 121-138. doi: 10.1007/s10665-011-9482-4
|
| [50] |
A. Oron, S. H. Davis, S. G. Bankoff, Long-scale evolution of thin liquid films, Rev. Mod. Phys., 69 (1997), 931-980. doi: 10.1103/RevModPhys.69.931
|
| [51] | T. G. Myers, Application of non-Newtonian models to thin film flow, Phys. Rev. E, 72 (2005), 066302. |
| [52] |
R. V. Craster, O. K. Matar, Dynamics and stability of thin liquid films, Rev. Mod. Phys., 81 (2009), 1131-1198. doi: 10.1103/RevModPhys.81.1131
|
| [53] |
P. E. King-Smith, C. G. Begley, R. J. Braun, Mechanisms, imaging and structure of tear film breakup, Ocul. Surf., 16 (2018), 4-30. doi: 10.1016/j.jtos.2017.09.007
|
| [54] |
K. L. Maki, R. J. Braun, P. Ucciferro, W. D. Henshaw, P. E. King-Smith, Tear film dynamics on an eye-shaped domain. Part 2. Flux boundary conditions, J. Fluid Mech., 647 (2010b), 361-390. doi: 10.1017/S002211200999382X
|
| [55] |
R. A. Luke, R. J. Braun, T. A. Driscoll, C. G. Begley, D. Awisi-Gyau, Parameter Estimation for
Evaporation-Driven Tear Film Thinning, Bull. Math. Biol., 82 (2020), 71. doi: 10.1007/s11538-020-00745-8
|
| [56] |
J. J. Nichols, G. L. Mitchell, P. E. King-Smith, Thinning rate of the precorneal and prelens tear films, Invest. Ophthalmol. Vis. Sci., 46 (2005), 2353-2361. doi: 10.1167/iovs.05-0094
|
| [57] |
R. A. Luke, R. J. Braun, T. A. Driscoll, D. Awisi-Gyau, C. G. Begley, Parameter estimation for mixed-mechanism tear film thinning, Bull. Math. Biol., 83 (2021), 1-39. doi: 10.1007/s11538-020-00834-8
|
| [58] | J. M. Tiffany, The viscosity of human tears, Int. Ophthalmol., 15 (1991), 371-376. |
| [59] |
B. Nagyová, J. M. Tiffany, Components of tears responsible for surface tension, Curr. Eye Res., 19 (1999), 4-11. doi: 10.1076/ceyr.19.1.4.5341
|
| [60] |
V. S. Ajaev, G. M. Homsy, Steady vapor bubbles in rectangular microchannels, J. Colloid Interface Sci., 240 (2001), 259-271. doi: 10.1006/jcis.2001.7562
|
| [61] |
Z. Wu, C. G. Begley, N. Port, A. Bradley, R. J. Braun, P. E. King-Smith, The effects of increasing ocular surface stimulation on blinking and tear secretion, Invest. Ophthalmol. Vis. Sci., 56 (2015), 4211-4220. doi: 10.1167/iovs.14-16313
|
| [62] | T. Casalini, M. Salvalaglio, G. Perale, M. Masi, C. Cavallotti, Diffusion and aggregation of sodium fluorescein in aqueous solutions, J. Phys. Chem. B, 115 (2011), 12896-12904. |
| [63] |
R. Riquelme, I. Lira, C. Pérez-López, J. A. Rayas, R. Rodríguez-Vera, Interferometric measurement of a diffusion coefficient: comparison of two methods and uncertainty analysis, J. Phys. D. Appl. Phys., 40 (2007), 2769-2776. doi: 10.1088/0022-3727/40/9/015
|
| [64] |
M. C. Mota, P. Carvalho, J. Ramalho, E. Leite, Spectrophotometric analysis of sodium fluorescein aqueous solutions. determination of molar absorption coefficient, Int. Ophthalmol., 15 (1991), 321-326. doi: 10.1007/BF00128951
|
| [65] |
K. L. Maki, R. J. Braun, W. D. Henshaw, P. E. King-Smith, Tear film dynamics on an eye-shaped domain I. pressure boundary conditions, Math. Med. Biol., 27 (2010a), 227-254. doi: 10.1093/imammb/dqp023
|
| [66] | L. Li, R. J. Braun, K. L. Maki, W. D. Henshaw, P. E. King-Smith, Tear film dynamics with evaporation, wetting and time-dependent flux boundary condition on an eye-shaped domain, Phys. Fluids, 26 (2014), 052101. |
| [67] |
L. Li, R. J. Braun, W. D. Henshaw, P. E. King-Smith, Computed flow and fluorescence over the ocular surface, Math. Med. Biol., 35 (2018), 51-85. doi: 10.1093/imammb/dqx011
|
| [68] |
A. A. V. Cruz, D. M. Garcia, C. T. Pinto, S. P. Cechetti, Spontaneous eyeblink activity, Ocul. Surf., 9 (2011), 29-41. doi: 10.1016/S1542-0124(11)70007-6
|
| [69] |
M. G. Doane, Interaction of eyelids and tears in corneal wetting and the dynamics of the normal human eyeblink, Am. J. Ophthalmol., 89 (1980), 507-516. doi: 10.1016/0002-9394(80)90058-6
|
| [70] |
T. A. Driscoll, R. J. Braun, J. K. Brosch, Simulation of parabolic flow on an eye-shaped domain with moving boundary, J. Eng. Math., 111 (2018), 111-126. doi: 10.1007/s10665-018-9957-7
|
| [71] | E. Aydemir, C. J. W. Breward, T. P. Witelski, The effect of polar lipids on tear film dynamics, Bull. Math. Biol., 73 (2010), 1171-1201. |
| [72] |
M. B. Jones, C. P. Please, D. L. S. McElwain, G. R. Fulford, A. P. Roberts, M. J. Collins, Dynamics of tear film deposition and draining, Math. Med. Biol., 22 (2005), 265-88. doi: 10.1093/imammb/dqi012
|
| [73] | M. B. Jones, D. L. S. McElwain, G. R. Fulford, M. J. Collins, and A. P. Roberts, The effect of the lipid layer on tear film behavior, Bull. Math. Biol., 68 (2006), 1355-81. |
| [74] |
M. Bruna, C. J. W. Breward, The influence of nonpolar lipids on tear film dynamics, J. Fluid Mech., 746 (2014), 565-605. doi: 10.1017/jfm.2014.106
|
| [75] |
A. Heryudono, R. J. Braun, T. A. Driscoll, L. P. Cook, K. L. Maki, P. E. King-Smith, Single-equation models for the tear film in a blink cycle: Realistic lid motion, Math. Med. Biol., 24 (2007), 347-377. doi: 10.1093/imammb/dqm004
|
| [76] |
Q. Deng, T. A. Driscoll, R. J. Braun, P. E. King-Smith, A model for the tear film and ocular surface temperature for partial blinks, Interfacial Phen. Ht. Trans., 1 (2013), 357-381. doi: 10.1615/InterfacPhenomHeatTransfer.v1.i4.40
|
| [77] | Q. Deng, R. J. Braun, T. A. Driscoll, Heat transfer and tear film dynamics over multiple blink cycles, Phys. Fluids, 26 (2014), 071901. |
| [78] |
V. S. Zubkov, C. J. Breward, E. A. Gaffney, Coupling fluid and solute dynamics within the ocular surface tear film: a modelling study of black line osmolarity, Bull. Math. Biol., 74 (2012), 2062-2093. doi: 10.1007/s11538-012-9746-9
|
| [79] | K. L. Maki, R. J. Braun, G. A. Barron, The influeance of a lipid reservoir on the tear film formation, Math. Med. Biol., 37 (2020), 364-389. |
| [80] |
V. S. Zubkov, C. J. W. Breward, E. A. Gaffney, Meniscal tear film fluid dynamics near marx's line, Bull. Math. Biol., 75 (2013), 1524-1543. doi: 10.1007/s11538-013-9858-x
|
| [81] |
P. E. King-Smith, C. G. Begley, R. J. Braun, Mechanisms, imaging and structure of tear film breakup, Ocul. Surf., 16 (2018), 4-30. doi: 10.1016/j.jtos.2017.09.007
|
| [82] |
P. E. King-Smith, K. S. Reuter, R. J. Braun, J. J. Nichols, K. K. Nichols, Tear film breakup and structure studied by simultaneous video recording of fluorescence and tear film lipid layer images, Invest. Ophthalmol. Vis. Sci., 54 (2013), 4900-4909. doi: 10.1167/iovs.13-11878
|
| [83] |
H. Owens, J. Phillips, Spreading of the tears after a blink: Velocity and stabilization time in healthy eyes, Cornea, 20 (2001), 484-487. doi: 10.1097/00003226-200107000-00009
|
| [84] | L. Zhong, C. F. Ketelaar, R. J. Braun, C. G. Begley, P. E. King-Smith, Mathematical modeling of glob-driven tear film breakup, Math. Med. Biol., (2018), 1-37. |
| [85] |
L. Zhong, R. J. Braun, C. G. Begley, P. E. King-Smith, Dynamics of fluorescent imaging for rapid tear thinning, Bull. Math. Biol., 81 (2019), 39-80. doi: 10.1007/s11538-018-0517-0
|
| [86] |
A. Sharma, E. Ruckenstein, Mechanism of tear film rupture and formation of dry spots on cornea, J. Coll. Interface Sci., 106 (1985), 12-27. doi: 10.1016/0021-9797(85)90375-3
|
| [87] |
A. Sharma, Energetics of corneal epithelial cell-ocular mucus-tear film interactions: Some surface-chemical pathways of corneal defense, Biophys. Chem., 47 (1993), 87-99. doi: 10.1016/0301-4622(93)80036-I
|
| [88] | E. Bitton, J. V. Lovasik, Longitudinal analysis of precorneal tear film rupture patterns, in Lacrimal Gland, Tear Film, and Dry Eye Syndromes 2, Boston, MA, 438 (1998), 381-389. |
| [89] |
N. Yokoi, G. A. Georgiev, Tear-film-oriented diagnosis for dry eye, Jpn. J. Ophthalmol., 63 (2019), 127-136. doi: 10.1007/s10384-018-00645-4
|
| [90] | J. N. Israelachvili, Intermolecular and Surface Forces, Academic press, San Diego San Francisco, New York, 2011. |
| [91] | R. J. Braun, T. A. Driscoll, C. G. Begley, P. E. King-Smith, J. I. Siddique, On tear film breakup (TBU): Dynamics and imaging, Math. Med. Biol., 45 (2018), 145-180. |
| [92] |
L. Li, R. J. Braun, A model for the human tear film with heating from within the eye, Phys. Fluids, 24 (2012), 062103. doi: 10.1063/1.4723870
|
| [93] |
K. L. Miller, K. A. Polse, C. J. Radke, Black line formation and the "perched" human tear film, Curr. Eye Res., 25 (2002), 155-162. doi: 10.1076/ceyr.25.3.155.13478
|
| [94] | L. N. Trefethen, Spectral Methods in MATLAB, SIAM, Philadelphia, 2000. |
| [95] | L. Zhornitskaya, A. L. Bertozzi, Positivity-preserving numerical schemes for lubrication-type equations, SIAM J. Numer. Anal., 37 (2000), 523-555. |
| [96] |
K. N. Winter, D. M. Anderson, R. J. Braun, A model for wetting and evaporation of a post-blink precorneal tear film, Math. Med. Biol., 27 (2010), 211-225. doi: 10.1093/imammb/dqp019
|
| [97] |
H. Liu, C. G. Begley, R. Chalmers, G. Wilson, S. P. Srinivas, J. A. Wilkinson, Temporal progression and spatial repeatability of tear breakup, Optom. Vis. Sci., 83 (2006), 723-730. doi: 10.1097/01.opx.0000237546.88464.6d
|
| [98] |
M. R. Stapf, R. J. Braun, P. E. King-Smith, Duplex tear film evaporation analysis, Bull. Math. Biol., 79 (2017), 2814-2846. doi: 10.1007/s11538-017-0351-9
|
| [99] | A. Sharma, E. Ruckenstein, The role of lipid abnormalities, aqueous and mucus deficiencies in the tear film breakup, and implications for tear substitutes and contact lens tolerance, J. Colloid Interface Sci., 111 (1986), 456-479. |
| [100] |
A. Sharma, E. Ruckenstein, Mechanism of tear film rupture and formation of dry spots on cornea, J. Colloid Interface Sci., 106 (1985), 12-27. doi: 10.1016/0021-9797(85)90375-3
|
| [101] | A. Sharma, E. Ruckenstein, An analytical nonlinear theory of thin film rupture and its application to wetting films, J. Coll. Interface Sci., 113 (1986), 8-34. |
| [102] |
L. Zhang, O. K. Matar, R. V. Craster, Analysis of tear film rupture: Effect of non-Newtonian rheology, J. Coll. Interface Sci., 262 (2003), 130-48. doi: 10.1016/S0021-9797(03)00200-5
|
| [103] |
L. Zhang, O. K. Matar, R. V. Craster, Rupture analysis of the corneal mucus layer of the tear film, Molec. Sim., 30 (2004), 167-72. doi: 10.1080/0892702031000152118
|
| [104] |
J. C. Pandit, B. Nagyová, A. J. Bron, J. M. Tiffany, Physical properties of stimulated and unstimulated tears, Exp. Eye Res., 68 (1999), 247-53. doi: 10.1006/exer.1998.0600
|
| [105] |
O. K. Matar, R. V. Craster, M. R. E. Warner, Surfactant transport on highly viscous surface films, J. Fluid Mech., 466 (2002), 85-111. doi: 10.1017/S0022112002001106
|
| [106] |
M. Dey, A. S. Vivek, H. N. Dixit, A. Richhariya, J. J. Feng, A model of tear-film breakup with continuous mucin concentration and viscosity profiles, J. Fluid Mech., 858 (2019), 352-376. doi: 10.1017/jfm.2018.776
|
| [107] | M. Dey, A. S. Vivek, H. N. Dixit, A. Richhariya, J. J. Feng, A model of tear-film breakup with continuous mucin concentration and viscosity profiles-corrigendum, J. Fluid Mech., 889 (2020), 1-2. |
| [108] |
P. E. King-Smith, E. A. Hinel, J. J. Nichols, Application of a novel interferometric method to investigate the relation between lipid layer thickness and tear film thinning, Invest. Ophthalmol. Vis. Sci., 51 (2010), 2418-2423. doi: 10.1167/iovs.09-4387
|
| [109] | L. Zhong, R. J. Braun, P. E. King-Smith, C. G. Begley, Mathematical modeling of glob-driven tear film breakup, J. Modeling Ophthalmol., 2 (2018), 24-28. |
| [110] |
C. G. Begley, T. L. Simpson, H. Liu, E. Salvo, Z. Wu, A. Bradley, et al., Quantitative analysis of tear film fluorescence and discomfort during tear film instability and thinning: Sensory responses during tear film breakup and thinning, Invest. Ophthalmol. Vis. Sci., 54 (2013), 2645-2653. doi: 10.1167/iovs.12-11299
|
| [111] | C. Belmonte, M. C. Acosta, J. Merayo-Lloves, J. Gallar, What causes eye pain?, Curr. Ophthalmol. Rep., 3 (2015), 111-121. |
| [112] |
D. Awisi-Gyau, C. G. Begley, P. Situ, T. L. Simpson, Changes in corneal detection thresholds after repeated tear film instability, Invest. Ophthalmol. Vis. Sci., 60 (2019), 4234. doi: 10.1167/iovs.19-27802
|
| [113] |
A. J. Bron, P. Argüeso, M. Irkec, F. V. Bright, Clinical staining of the ocular surface: Mechanisms and interpretations, Prog. Ret. Eye Res., 44 (2015), 36-61. doi: 10.1016/j.preteyeres.2014.10.001
|
| [114] |
A. Sharma, Acid-base interactions in the cornea-tear film system: surface chemistry of corneal wetting, cleaning, lubrication, hydration and defense, J. Dispersion Sci. Technol., 19 (1998), 1031-1068. doi: 10.1080/01932699808913229
|
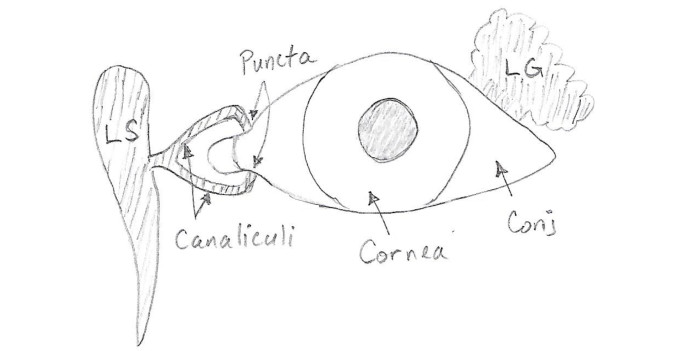
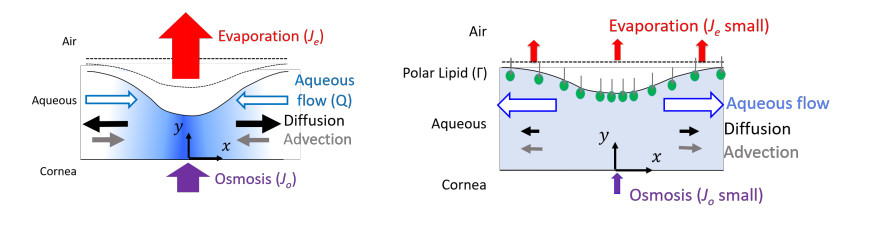
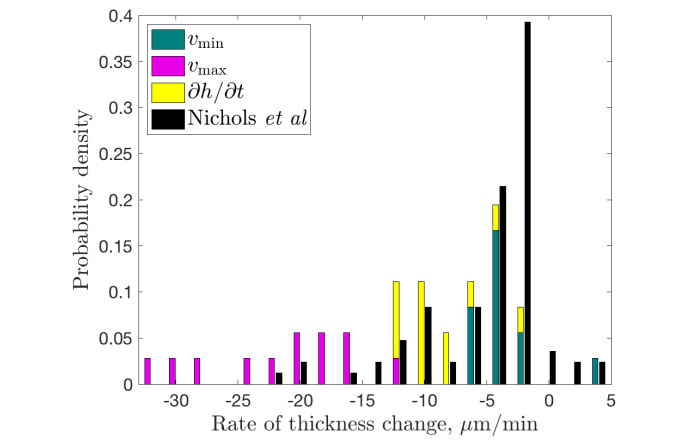

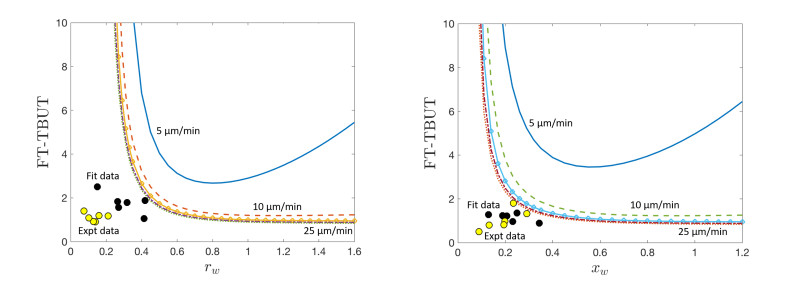
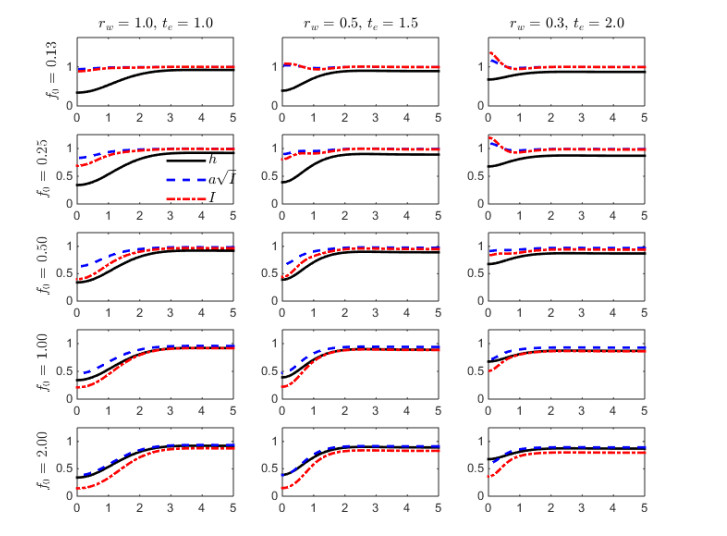
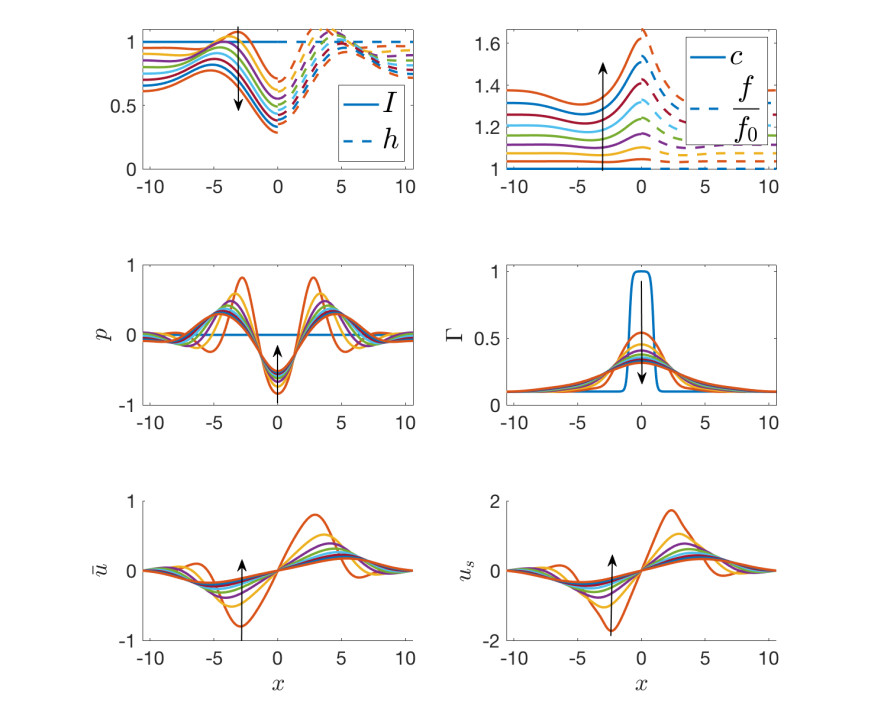
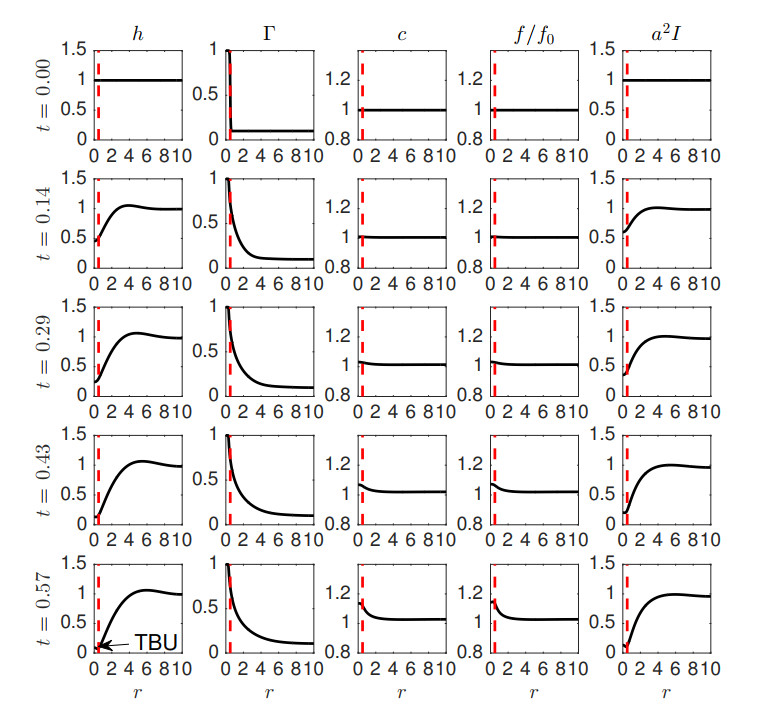
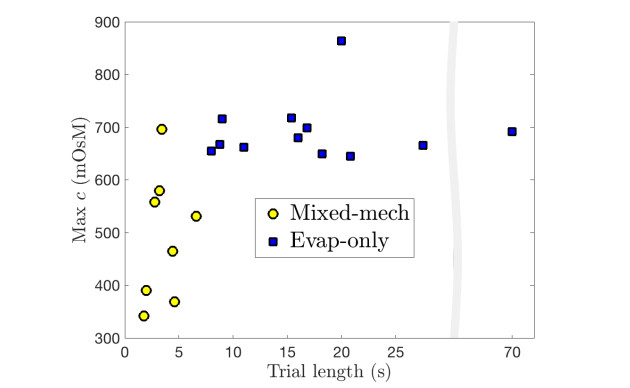
Figures(9) / Tables(3)

Richard J. Braun, Rayanne A. Luke, Tobin A. Driscoll, Carolyn G. Begley. Dynamics and mechanisms for tear breakup (TBU) on the ocular surface[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5146-5175. doi: 10.3934/mbe.2021262







 DownLoad:
DownLoad: