Citation: Eun-jin Kim, Massimo Capoccia. Mechano-electric effect and a heart assist device in the synergistic model of cardiac function[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5212-5233. doi: 10.3934/mbe.2020282

| [1] | H. Haken, Information and Self-Organization: A Macroscopic Approach to Complex Systems, Springer Series in Synergetics, Heidelberg: Springer-Verlag, Berlin, 2006. |
| [2] | T. Wright, J. Twaddle, C. Humphries, S. Hayes, E. Kim, Variability and degradation of selforganization in self-sustained oscillators, Math. Biosci., 273 (2016), 57-69. |
| [3] | K. Ascrof, Available from: https://sheffield.academia.edu/KieranAscroft (accessed on 23 August 2019). |
| [4] | Z. Knudsen, A. Holden, J. Brindley, Qualitative modeling of mechano-electrical feedback in a ventricular cell, Bull. Math. Biol., 6 (1997), 115-181. |
| [5] | G. Tse, S. T. Wong, V. Tse, Y. T. Lee, H. Y. Lin, J. M. Yeo, Cardiac dynamics: Alternans and arrhythmogenesis, J. Arrhythm, 32 (2016), 411-417. |
| [6] | M. R. Franz, Mechano-electrical feedback in ventricular myocardium, Cardiovascular Res., 32 (1996), 15-24. |
| [7] | M. J. Lab, D. G. Allen, C. H. Orchard, The effects of shortening on myoplasmic calcium concentration and action potential in mammalian ventricular muscle, Circ. Res., 55 (1984), 825- 829. |
| [8] | A. Kamkin, I. Kiseleva, K. D. Wagner, H. Scholz, Mechano-Electric Feedback in the Heart: Evidence from Intracellular Microelectrode Recordings on Multicellular Preparations and Single Cells from Healthy and Diseased Tissue, in Mechanosensitivity in Cells and Tissues (eds. A. Kamkin and I. Kiseleva), Moscow: Academia, 2005. |
| [9] | P. Kohl, P. Hunter, D. Noble, Stretch-induced changes in heart rate and rhythm: clinical observations, experiments and mathematical models, in Biophysics and Molecular Biology, 71 (1999), 91-138. |
| [10] | A. Collet, J. Bragard, P. C. Dauby, Temperature, geometry, and bifurcations in the numerical modeling of the cardiac mechano-electric feedback, Chaos, 27 (2017), 093924. |
| [11] | S. Galice, D. M. Bers, D. Sato, Stretch-activated current can promote or suppress cardiac alternans depending on voltage-calcium interaction, Biophys. J., 110 (2016), 2671-2677. |
| [12] | O. Bikou, C. Kho, K. Ishikawa, Atrial stretch and arrhythmia after myocardial infarction, Aging, 11 (2019), 11-12. |
| [13] | P. Kohl, A. D. Nesbitt, P. J. Cooper, M. Lei, Sudden cardiac death by commotio cordis: role of mechano-electric feedback, Cardiovasc. Res., 50 (2001), 280-289. |
| [14] | C. Madias, M. B. J. J. Weinstock, N. R. Estes, M. S. Link, Commotio cordis-sudden cardiac death with chest wall impact, J. Cardiovasc. Electrophysiol., 18 (2007), 115-122. |
| [15] | W. Li, P. Kohl, N. Trayanova, Induction of ventricular arrhythmias following mechanical impact: a simulation study in 3d, J. Mol. Histol., 35 (2004), 679-686. |
| [16] | W. Li, P. Kohl, N. Trayanova, Myocardial ischemia lowers precordial thump ef!cacy: an inquiry into mechanisms using three-dimensional simulations, Heart Rhythm, 3 (2006), 179-186. |
| [17] | M. J. Lab, Transient depolarization and action potential alterations following mechanical changes in isolated myocardium, Cardiovasc. Res., 14 (1980), 624-637. |
| [18] | M. R. Franz, D. Burkho, D. T. Yue, K. Sagawa, Mechanically induced action potential changes and arrhythmia in isolated and in situ canine hearts, Cardiovasc. Res., 23 (1989), 213-223. |
| [19] | H. Hu, F. Sachs, Stretch-induced ion channels in the heart, J. Mol. Cell. Cardiol., 29 (1997), 1511-1523. |
| [20] | E. G. Tolkacheva, D. G. Schaeffer, D. J. Gauthier, C. C. Mitchell, Analysis of the Fenton-Karma model through an approximation by a one-dimensional map, Chaos, 12 (2002), 1034-1042. |
| [21] | M. Radszuweit, E. Alvarez-Lacalle, M. Bar, B. Echebarria, Cardiac contraction induces discordant alternans and localized block, Phys. Rev. E, 91 (2015), 022703. |
| [22] | A. Hazim, Y. Belhamadia, S. Dubljevicet, Effects of mechano-electrical feedback on the onset of alternans: A computational study, Chaos, 29 (2019), 063126. |
| [23] | F. Fenton, A. Karma, Vortex dynamics in three-dimensional continuous myocardium with fiber rotation: Filament instability and fibrillation, Chaos, 8 (1998), 20-47. |
| [24] | F. Yapari, D. Deshpande, Y. Belhamadia, S. Dubljevic, Control of cardiac alternans by mechanical and electrical feedback, Phys. Rev. E, 90 (2014), 012706. |
| [25] | L. D. Weise, A. V. Panfilov, Discrete Mechanical Modeling of Mechanoelectrical Feedback in Cardiac Tissue: Novel Mechanisms of Spiral Wave Initiation, in Modeling the Heart and the Circulatory System, Springer, Cham, 14 (2015), 29-50. |
| [26] | A. Hazim, Y. Belhamadia, S. Dubljevicet, Control of cardiac alternans based on electromechanical model for cardiac tissue, Comput. Biol. Med., 63 (2015), 108117. |
| [27] | C. Franzone, L. F. Pavarino, S. Scacchiet, Effects of mechanical feedback on the stability of cardiac scroll waves: A bidomain electro-mechanical simulation study, Chaos, 27 (2017), 093905. |
| [28] | A. Hazim, Y. Belhamadia, S. Dubljevic, Mechanical perturbation control of cardiac alternans, Phys. Rev. E, 97 (2018), 052407. |
| [29] | A. Amar, S. Zlochiver, O. Barnea, Mechano-electric feedback effects in a three-dimensional (3D) model of the contracting cardiac ventricle, PLoSONE, 13 (2018), e0191238. |
| [30] | R. R. Alievand, A. V. Panfilov, A simple two-variable model of cardiac excitation, Chaos, Solitons Fractals, 7 (1996), 293-301. |
| [31] | A. Bueno-Orovio, E. M. Cherry, H. Flavio, F. H. Fenton, Minimal model for human ventricular action potentials in tissue, J. Theo. Biology, 253 (2008), 544-560. |
| [32] | V. Timmermann, L. A. Dejgaard, K. H. Haugaa, A. G. Edwards, J. Sundnes, A. D. McCulloch, et al., An integrative appraisal of mechano-electric feedback mechanisms in the heart, Prog. Biophys. Mol. Biol., 130 (2017), 404-417. |
| [33] | M. Varela, A. Roy, J. Lee, A survey of pathways for mechano-electric coupling in the Atria, preprint, arXiv:2005.08121. |
| [34] | Z. Qu, G. Hu, A. Garfinkel, J. N. Weiss, Nonlinear and stochastic dynamics in the heart, Phys. Rep., 543 (2014), 61-162. |
| [35] | A. P. Newton, E. Kim, On the self-organizing process of large scale shear flows, Phys. Plasmas, 20 (2013), 092306. |
| [36] | H. Suga, K. Sagawa, A. A. Shoukas, Load independence of the instantaneous pressure-volume ratio of the canine left ventricle and effects of epinephrine and heart rate on the ratio, Circ. Res., 32 (1973), 314-322. |
| [37] | K. Sagawa, The ventricular pressure-volume diagram revisited, Circ. Res., 43 (1978), 677-687. |
| [38] | M. A. Simaan, Rotary heart assist devices, in Springer Handbook of Automation, Springer, (2009), 1409-1421. |
| [39] | M. A. Simaan, A. Ferreira, S. Chen, J. F. Antaki, D. G. Galati, A dynamical state space representation and performance analysis of feedback-controlled rotary left ventricular assist device IEEE Trans, Control Syst. Tech., 17 (2019), 15-28. |
| [40] | T. E. Claessens, D. Georgakopoulos, M. Afanasyeva, S. J. Vermeersch, H. D. Millar, N. Stergiopulos, et al., Nonlinear isochrones in murine left ventricular pressure-volume loops: how well does the time-varying elastance concept hold?, Am. J. Physiol. Heart Circ. Physiol., 290 (2006), H1474-H1483. |
| [41] | S. Vandenberghe, P. Segers, P. Steendijk, B. Meyns, R. A. E. Dion, J. F. Antaki, et al., Modelling ventricular function during cardiac assist: Does time-varying elastance work?, Am. Soc. Artif. Intern. Organs J., 52 (2006), 4-8. |
| [42] | E. Kim, M. Capoccia, Synergistic model of cardiac function with a heart assist device, Bioengineering, 7 (2019), 1-16. |
| [43] | M. Capoccia, Development and characterization of the arterial windkessel and its role during left ventricular assist device assistance, Artif. Organs, 39 (2015), E138. |
| [44] | M. Capoccia, S. Marconi, S. A. Singh, D. M. Pisanelli, C. De Lazzari, Simulation as a preoperative planning approach in advanced heart failure patients: A retrospective clinical analysis, Bio. Med. Eng. OnLine, 7 (2018), 52. |
| [45] | G. Ferrari, A. Di Molfetta, K. Zieliński, L. Fresiello, Circulatory modelling as a clinical decision support and an educational tool, Biomed. Data J., 1 (2015), 45-50. |
| [46] | J. Bestel, F. Clément, M. Sorine, A biomechanical model of muscle contraction, in Medical Image Computing and Computer-Assisted Intervention MICCAI 2001, Springer, Berlin, Heidelberg, (2001), 1159-1161. |
| [47] | J. Bestel, Modèle différentiel de la contraction musculaire contrôlée: Application au système cardio-vasculaire, Ph.D thesis, Universit Paris 9, 2000. |
| [48] | Available from: https://www.cellml.org/ (accessed on 8 July 2020). |
| [49] | Available from: https://www.scholarpedia.org/article/Models of cardiac cell (accessed on 8 July 2020). |
| [50] | E. Ott, Chaos in Dynamical Systems, Cambridge University Press, 1993. |
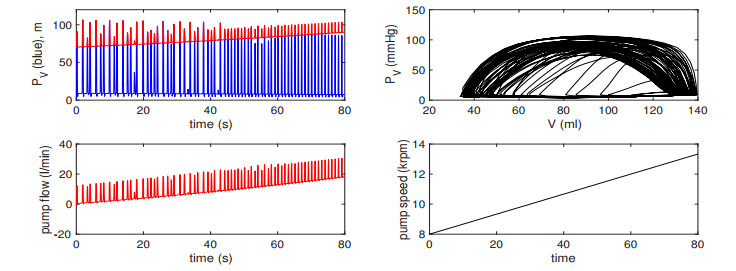
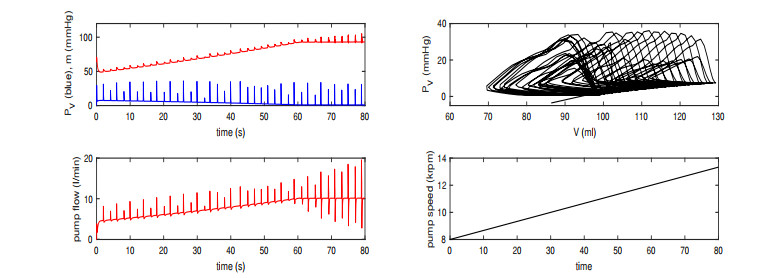
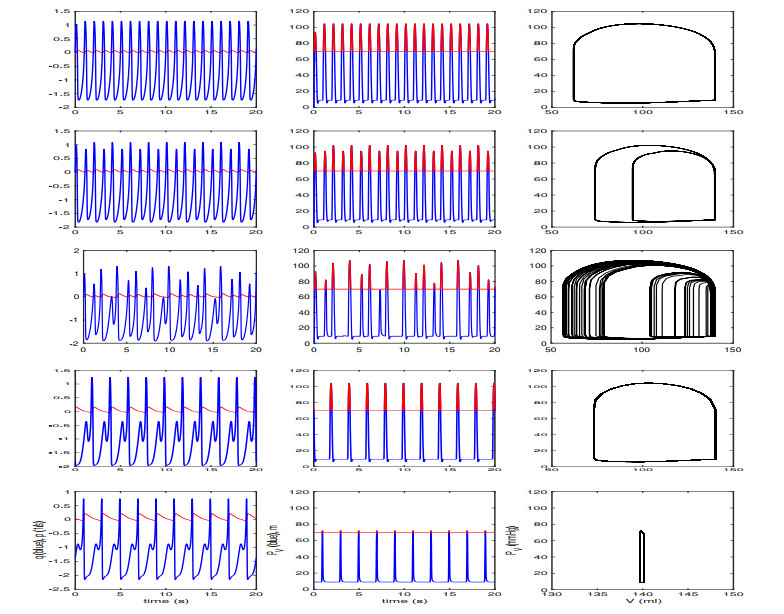
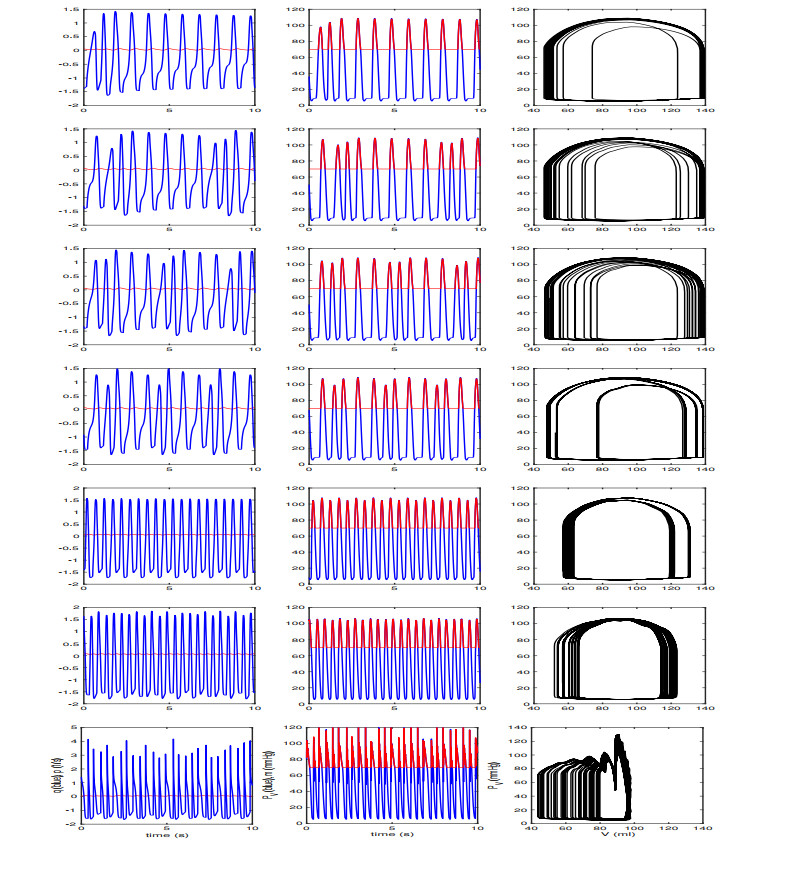
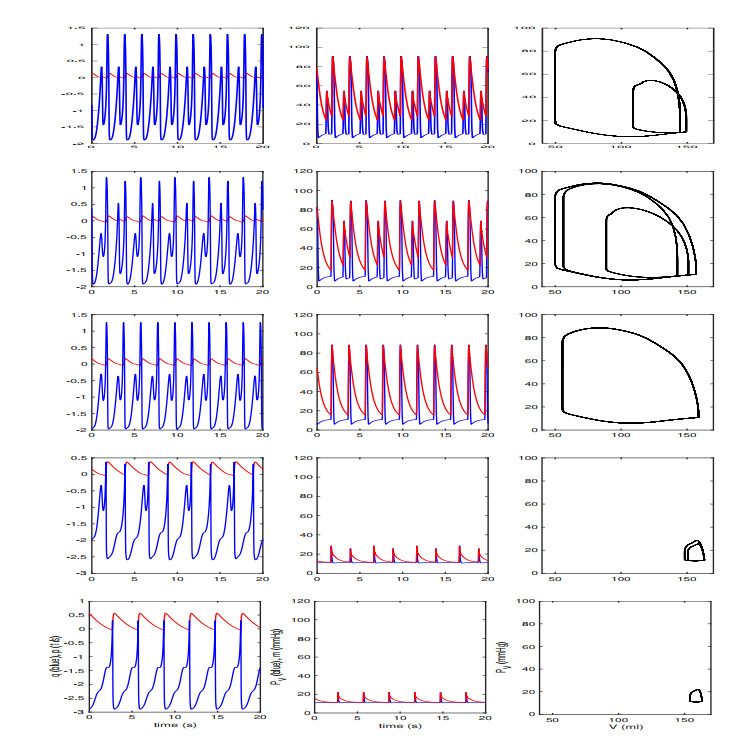
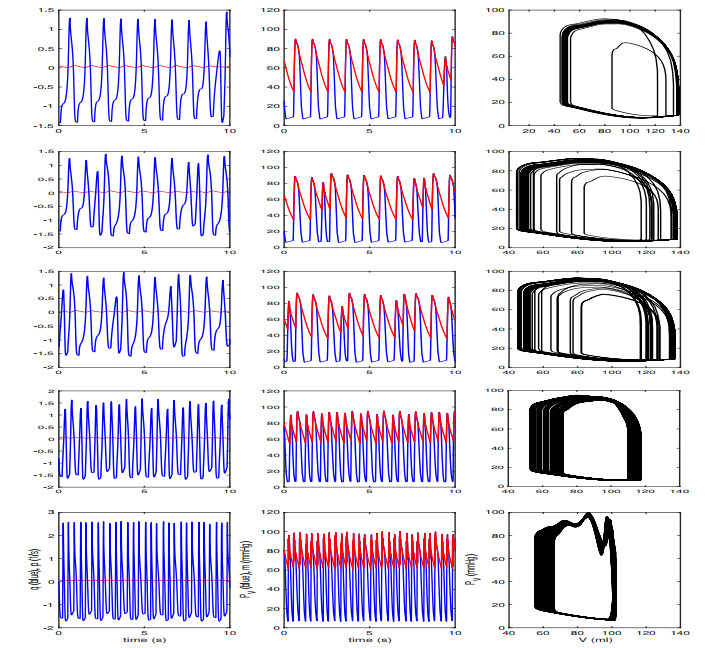
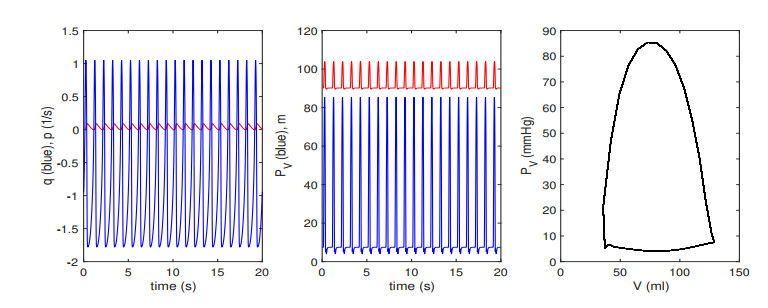
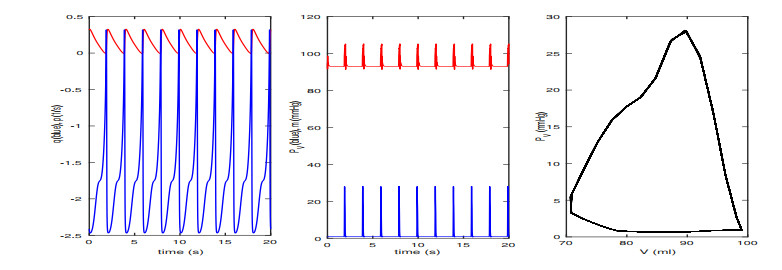
Figures(13) / Tables(2)

Eun-jin Kim, Massimo Capoccia. Mechano-electric effect and a heart assist device in the synergistic model of cardiac function[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5212-5233. doi: 10.3934/mbe.2020282







 DownLoad:
DownLoad: