Citation: Hua Wang, Weiwei Li, Wei Huang, Jiqiang Niu, Ke Nie. Research on land use classification of hyperspectral images based on multiscale superpixels[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5099-5119. doi: 10.3934/mbe.2020275

| [1] | F. Yulianto, Suwarsono, U. C. Nugroho, N. P. Nugroho, W. Sunarmodo, M. R. Khomarudin, Spatial-Temporal Dynamics Land Use/Land Cover Change and Flood Hazard Mapping in the Upstream Citarum Watershed, West Java, Indonesia, Quaestiones Geographicae, 39 (2020), 125-146. |
| [2] | Y. Ye, Y. An, B. Chen, J. Wang, Y. Zhong, Land use classification from social media data and satellite imagery, J. Supercomput., 76 (2020), 777-792. |
| [3] | A. F. H. Goetz, Three decades of hyperspectral remote sensing of the Earth: A personal view, Remote Sens. Environ., 113 (2009), S5-S16. |
| [4] | L Sun, C Ma, Y Chen, Y. Zheng, H. J. Shim, Z. Wu, Low Rank Component Induced Spatial-spectral Kernel Method for Hyperspectral Image Classification, IEEE Trans. Circuits Syst. Video Technol., 2019 (2019), 3005-3008. |
| [5] | L Sun, F Wu, T Zhan, W. Liu, J. Wang, B. Jeon, Weighted Nonlocal Low-Rank Tensor Decomposition Method for Sparse Unmixing of Hyperspectral Images, IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens., 13 (2020), 1174-1188. |
| [6] | J. Peng, L. Li, Y. Y. Tang, Maximum Likelihood Estimation-Based Joint Sparse Representation for the Classification of Hyperspectral Remote Sensing Images, IEEE Trans. Neural Networks Learn. Syst., 30 (2018), 1790-1802. |
| [7] | J. C. W. Chan, D. Paelinckx, Evaluation of Random Forest and Adaboost tree-based ensemble classification and spectral band selection for ecotope mapping using airborne hyperspectral imagery, Remote Sens. Environ., 112 (2008), 2999-3011. |
| [8] | P. Rao, J. Wang, Y. Wang, Extraction of information on construction land based on multi-feature decision tree classification, Trans. Chin. Soc. Agric. Eng., 30 (2104), 233-240. |
| [9] | Y. Liu, L. Wang, B. Zhang, J. Men, Scene-level land use classification based on multi-features soft-probability cascading, Trans. Chin. Soc. Agric. Eng., 32 (2016), 266-272. |
| [10] | K. Rangzan, M. Kabolizadeh, D. Karimi, S. Zareie, Supervised cross-fusion method: A new triplet approach to fuse thermal, radar, and optical satellite data for land use classification, Environ. Monit. Assess., 191 (2019), 481. |
| [11] | S. A. Manaf, N. Mustapha, M. N. Sulaiman, N. A. Husin, M. R. A. Hamid, Artificial Neural Networks for Satellite Image Classification of Shoreline Extraction for Land and Water Classes of the North West Coast of Peninsular Malaysia, Adv. Sci. Lett., 24 (2018), 1382-1387. |
| [12] | J. Fan, T. Chen, S. Lu, Unsupervised Feature Learning for Land-Use Scene Recognition, IEEE Trans. Geosci. Remote Sens., 55 (2017), 2250-2261. |
| [13] | J. Men, L. Fang, Y. Liu, Y. Sun, Land Use Classification Based on Multi-structure Convolution Neural Network Features Cascading, Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci., 2019 (2019), 163-167. |
| [14] | K. Bhosle, V. Musande, Evaluation of Deep Learning CNN Model for Land Use Land Cover Classification and Crop Identification Using Hyperspectral Remote Sensing Images, J. Indian Soc. Remote Sens., 47 (2019), 1949-1958. |
| [15] | S. Bera, V. K. Shrivastava, Analysis of various optimizers on deep convolutional neural network model in the application of hyperspectral remote sensing image classification, Int. J. Remote Sens., 41 (2020), 2664-2683. |
| [16] | W. Huang, Y. Xu, X. Hu, Z. Wei, Compressive Hyperspectral Image Reconstruction Based on Spatial-Spectral Residual Dense Network, IEEE Geosci. Remote Sens. Lett., 17 (2020), 884-888. |
| [17] | P. J. Du, J. S. Xia, Z. H. Xue, K. Tan, H. J. Su, R. Bao, Review of hyperspectral remote sensing image classification, J. Remote Sens., 20 (2016), 236-256. |
| [18] | J. Fan, T. Chen, S. Lu, Superpixel Guided Deep-Sparse-Representation Learning for Hyperspectral Image Classification, IEEE Trans. Circuits Syst. Video Technol., 28 (2018), 3163-3173. |
| [19] | J. Li, J. M. Bioucas-Dias, A. Plaza, Semisupervised Hyperspectral Image Segmentation Using Multinomial Logistic Regression With Active Learning, IEEE Trans. Geosci. Remote Sens., 48 (2010), 4085-4098. |
| [20] | P. Ghamisi, M. S. Couceiro, F. M. Martins, J. A. Benediktsson, Multilevel Image Segmentation Based on Fractional-Order Darwinian Particle Swarm Optimization, IEEE Trans. Geosci. Remote Sens., 52 (2014), 2382-2394. |
| [21] | P. Y. Wang, H. Q. Zhu, N. CHEN, UMMS: Efficient Superpixel Segmentation Driven by a Mixture of Spatially Constrained Uniform Distribution, IEICE Trans. Inf. Syst., 103 (2020), 181-185. |
| [22] | D. Song, X. Tan, B. Wang, L. Zhang, X. Shan, J. Cui, Integration of super-pixel segmentation and deep-learning methods for evaluating earthquake-damaged buildings using single-phase remote sensing imagery, Int. J. Remote Sens., 41 (2020), 1040-1066. |
| [23] | B. Amin, M. M. Riaz, A. Ghafoor, Automatic Image Matting of Synthetic Aperture Radar Target Chips, Radioengineering, 29 (2020), 228-234. |
| [24] | X. Yuan, S. Guo, C. Li, B. Lu, S. Lou, Near Infrared Star Centroid Detection by Area Analysis of Multi-Scale Super Pixel Saliency Fusion Map, Tsinghua Sci. Technol., 24 (2019), 291-300. |
| [25] | K. Tang, Z. Su, W. Jiang, J. Zhang, Superpixels for large dataset subspace clustering, Neural Comput. Appl., 31 (2019), 8727-8736. |
| [26] | Y. J. Chen, C. Y. Ma, Edge-Modified Superpixel Based Spectral-Spatial Kernel Method for Hyperspectral Image Classification, Acta Electron. Sin., 47 (2019), 73-81. |
| [27] | Z. Liu, Y. Wu, Y. Zou, Multiscale infrared superpixel-image model for small-target detection, J. Image Graphics, 24 (2019), 2159-2173. |
| [28] | M. S. Chaibou, P. H. Conze, K. Kalti, M. A. Mahjoub, B. Solaiman, Learning contextual superpixel similarity for consistent image segmentation, Multimedia Tools Appl., 79 (2020), 2601-2627. |
| [29] | Y. Song, W. Liu, S. Zong, Y. Luo, Segmentation Algorithm for Unmanned Aerial Vehicle Imagery Based on Superpixel and Ultrametric Contour Map, J. Comput. Aided Des. Comput. Graphics, 31 (2019), 1294-1300. |
| [30] | X. Hou, H. Zhao, Y. Ma, Fast Image Segmentation Algorithm Based on Superpixel Multi-feature Fusion, Acta Electron. Sin., 47 (2019), 2126-2133. |
| [31] | H. Liang, B. X. Yao, P. D. Chen, Superpixel segmentation method of high resolution remote sensing images based on hierarchical clustering, J. Infrared Millimeter Waves, 39 (2020), 263-272. |
| [32] | Z. Yang, X. D. Mu, S. Y. Wang, C. H. Ma, Scene classification of remote sensing images based on multiscale features fusion, Opt. Precis. Eng., 26 (2018), 3099-3107. |
| [33] | D. Zhang, T. Yin, G. Yang, M. Xia, L. Li, X. Sun, Detecting image seam carving with low scaling ratio using multi-scale spatial and spectral entropies, J. Visual Commun. Image Representation, 48 (2017), 281-291. |
| [34] | Z. Chen, X. Wang, K. Yan, J. Zheng, Deep multi-scale feature fusion for pancreas segmentation from CT images, Int. J. Comput. Assisted Radiol. Surg., 15 (2020), 415-423. |
| [35] | Hyperspectral Image, the National Mall in Washington DC. Available from: https://engineering.purdue.edu/~biehl/MultiSpec/hyperspectral.html. |
| [36] | M Graña, M. A Veganzons, B Ayerdi, Hyperspectral Remote Sensing Scenes. Available from: http://www.ehu.eus/ccwintco/index.php?title=Hyperspectral_Remote_Sensing_Scenes. |
| [37] | X. Jia, J. A. Richards, Segmented principal components transformation for efficient hyperspectral remote-sensing image display and classification, IEEE Trans. Geosci. Remote Sens., 37 (1999), 538-542. |
| [38] | X. Kang, P. Duan, S. Li, Hyperspectral image visualization with edge-preserving filtering and principal component analysis, Inf. Fusion, 57 (2020), 130-143. |
| [39] | B. Fatima, A. R. Shahid, S. Ziauddin, A. A. Safi, H. Ramzan, Driver Fatigue Detection Using Viola Jones and Principal Component Analysis, Appl. Artif. Intell., 34 (2020), 456-483. |
| [40] | Z. Zhang, J. Liu, Z. Xi, A disparity optimization algorithm using entropy rate super-pixel segmentation consistency check, Comput. Eng. Appl., 56 (2020), 1-8. |
| [41] | Liu Xia, Guo Yanan, Remote sensing image change detection algorithm based on random forest, Bull. Surv. Mapp., 5 (2020), 16-20. |
| [42] | C. Shi, C. M. Pun, Multiscale Superpixel-Based Hyperspectral Image Classification Using Recurrent Neural Networks With Stacked Autoencoders, IEEE Trans. Multimedia, 22 (2020), 487-501. |
| [43] | C. G. Karydas, Optimization of multi-scale segmentation of satellite imagery using fractal geometry, Int. J. Remote Sens., 41 (2020), 2905-2933. |
| [44] | L Sun, C Ma, Y Chen, H. J. Shim, Z. Wu, B. Jeon, Adjacent superpixel-based multiscale spatial-spectral kernel for hyperspectral classification, IEEE J. Selected Topics in Applied Earth Observations Remote Sens., 12 (2019), 1905-1919. |
| [45] | C. Liu, L. Hong, J. Chen, S. S. Chun, M. Deng, Fusion of pixel-based and multi-scale region-based features for the classification of high-resolution remote sensing image, J. Remote Sens., 19 (2015), 228-239. |
| [46] | G. Camps-Valls, L. Bruzzone, Kernel-based methods for hyperspectral image classification, IEEE Trans. Geosci. Remote Sens., 43 (2005), 1351-1362. |
| [47] | Y. Chen, J. Wang, R. Xia, Q. Zhang, Z. Cao, K. Yang, The Visual Object Tracking Algorithm Research Based on Adaptive Combination Kernel, J. Ambient Intell. Humanized Comput., 10 (2019), 4855-4867. |
| [48] | Y. Chen, J. Xiong, W. Xu, J. Zuo, A novel online incremental and decremental learning algorithm based on variable support vector machine, Cluster Comput., 22 (2019), 7435-7445. |
| [49] | Y. Chen, W. Xu, J. Zuo, K. Yang, The fire recognition algorithm using dynamic feature fusion and IV-SVM classifier, Cluster Comput., 22 (2019), 7665-7675. |
| [50] | K. Shankar, S. K. Lakshmanaprabu, D. Gupta, A. Maseleno, V. Hugo, C. de Albuquerque, Optimal feature-based multi-kernel SVM approach for thyroid disease classification, J. Supercomput., 76 (2020), 1128-1143. |
| [51] | M. Ramzan, A. Abid, H. U. Khan, S. M. Awan, A. Ismail, M. Ahmed, et al., A Review on State-of-the-Art Violence Detection Techniques, IEEE Access, 7 (2019), 107560-107575. |
| [52] | C. C. Chang, H. T. Huang, Automatic Tuning of the RBF Kernel Parameter for Batch-Mode Active Learning Algorithms: A Scalable Framework, IEEE Trans. Cybern., 49 (2019), 4460-4472. |
| [53] | K. Shang, P. Li, T. Cheng, Land Cover Classification of Hyperspectral Data Using Composite Kernel Support Vector Machines, Acta Sci. Nat. Univ. Pekin., 7 (2019), 107560-107575. |

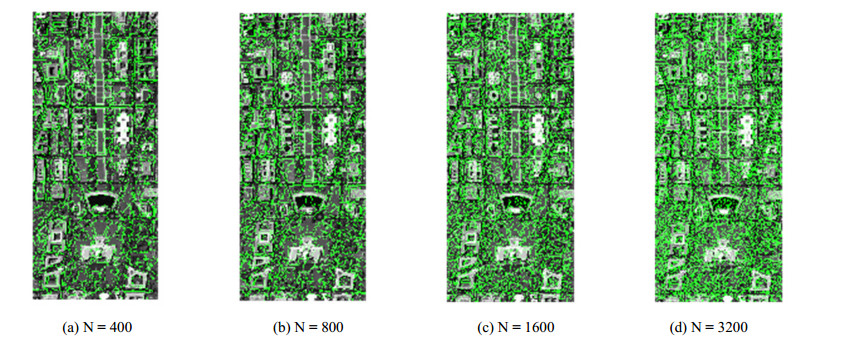
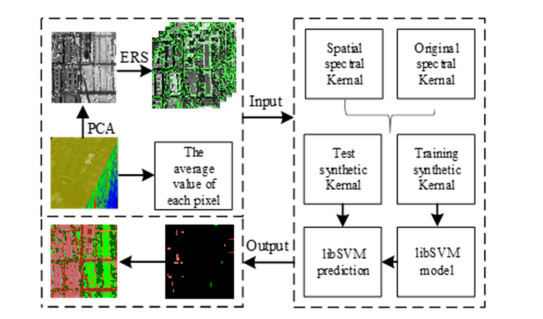
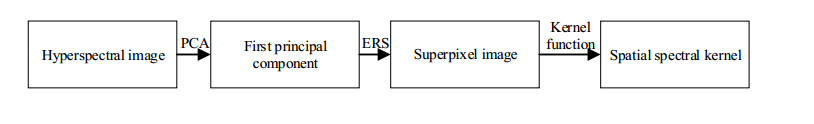
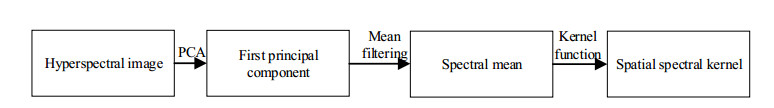

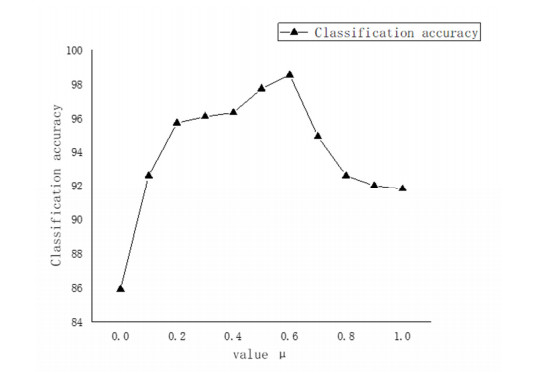



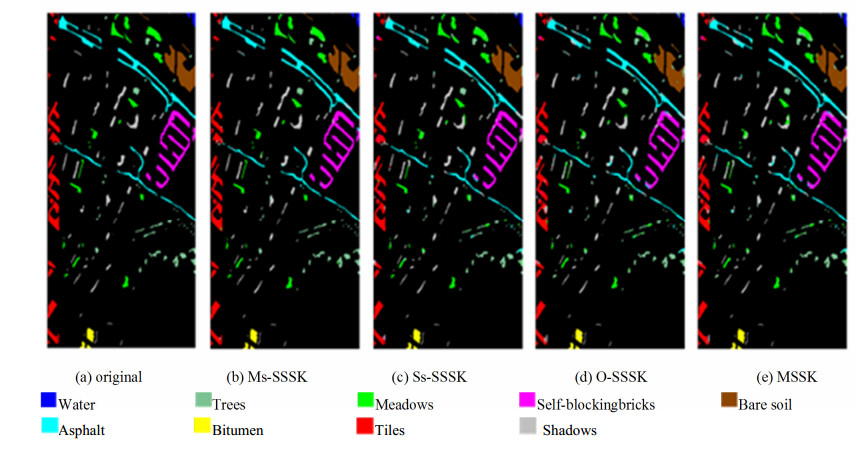
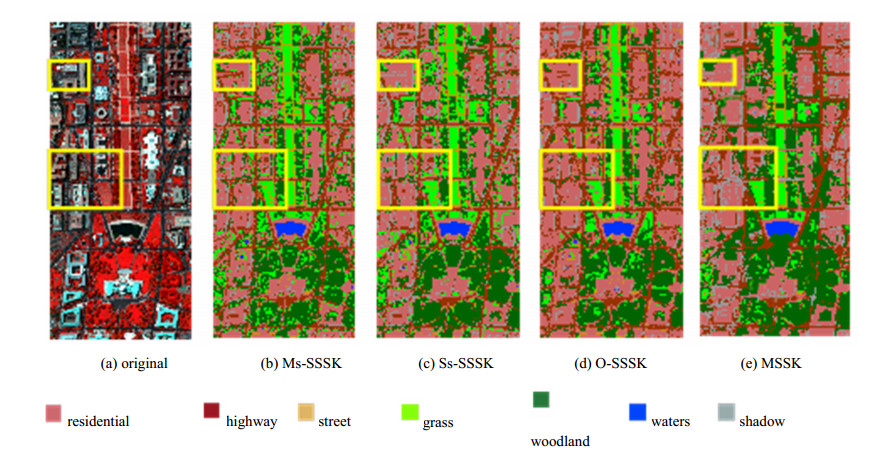
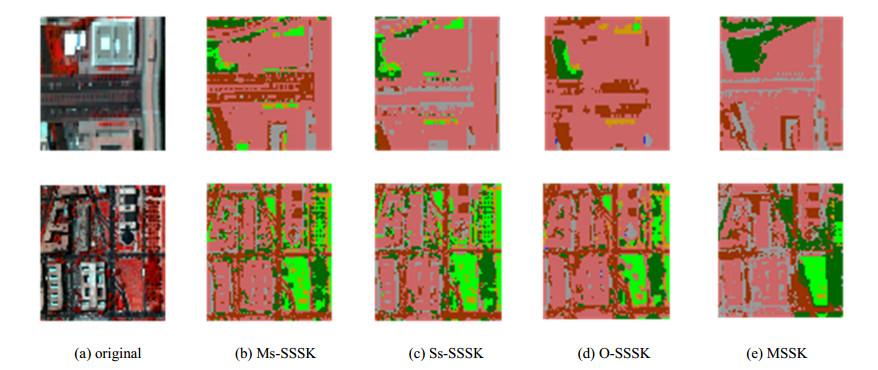
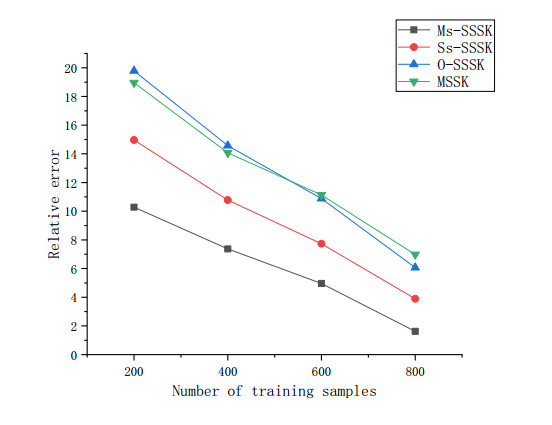
Figures(14) / Tables(3)

Hua Wang, Weiwei Li, Wei Huang, Jiqiang Niu, Ke Nie. Research on land use classification of hyperspectral images based on multiscale superpixels[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5099-5119. doi: 10.3934/mbe.2020275







 DownLoad:
DownLoad: