Citation: Xu Bie, Yuanyuan Tang, Ming Zhao, Yingxi Liu, Shen Yu, Dong Sun, Jing Liu, Ying Wang, Jianing Zhang, Xiuzhen Sun. Pilot study of pressure-flow properties in a numerical model of the middle ear[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2418-2431. doi: 10.3934/mbe.2020131

| [1] | K. J. Kvaerner, P. Nafstad, J. J. K. Jaakkola, Upper respiratory morbidity in preschool children: A cross-sectional study, Arch. Otolaryngol. Head Neck Surg., 126 (2000), 1201-1206. |
| [2] | C. D. Bluestone, J. O. Klein, Physiology, pathophysiology and pathogenesis, Otitis Media Infants Child., 2 (2007), 17-38. |
| [3] | R. W. Ranakusuma, Y. Pitoyo, E. D. Safitri, S. Thorning, E. M. Beller, S. Sastroasmoro, et al., Systemic corticosteroids for acute otitis media in children, Cochrane Database Syst. Rev., 3 (2018), CD012289. |
| [4] | D. S. Hurst, The role of allergy in otitis media with effusion, Otolaryngol. Clin. North Am., 44 (2011), 637-654. |
| [5] | L. H. Nguyen, J. J. Manoukian, S. E. Sobol, D. S. Melvin, Similar allergic inflammation in the middle ear and the upper airway: evidence linking otitis media with effusion to the united airways concept, J. Allergy Clin. Immunol., 114 (2004), 1110-1115. |
| [6] | M. E. Smith, J. R. Tysome, Tests of Eustachian tube function: a review, Clin. Otolaryngol., 40 (2015), 300-311. |
| [7] | M. E. Smith, D. J. Scoffings, J. R. Tysome, Imaging of the Eustachian tube and its function: A systematic review, Neuroradiology, 58 (2016), 543-556. |
| [8] | A. Teymoortash, S. Hamzei, T. Murthum, B. Eivazi, I. Kureck, J. A. Werner, Temporal bone imaging using digital volume tomography and computed tomography: a comparative cadaveric radiological study, Surg. Radiol. Anat., 33 (2011), 123-128. |
| [9] | A. Morra, G. Tirelli, A. Rimondini, V. Cioffi, M. Russolo, V. Giacomarra, et al., Usefulness of virtual endoscopic three-dimensional reconstructions of the middle ear, Acta Otolaryngol., 122 (2002), 382-385. |
| [10] | Z. Csakanyi, G. Katona, E. Josvai, F. Mohos, I. Sziklai, Volume and surface of the mastoid cell system in otitis media with effusion in children: A case-control study by three-dimensional reconstruction of computed tomographic images, Otol. Neurotol., 32 (2011), 64-70. |
| [11] | J. T. Vrabec, S. W. Champion, J. D. Gomez, R. J. Johnson, G. Chaljub, 3D CT imaging method for measuring temporal bone aeration, Acta Otolaryngol., 122 (2002), 831-835. |
| [12] | S. Yu, X. Sun, Y. Liu, Numerical Analysis of the Relationship between Nasal Structure and Its Function, Sci. World J., 2014 (2014),1-6. |
| [13] | X. Sun, C. Yu, Y. Wang, Y. Liu, Numerical simulation of soft palate movement and airflow in human upper airway by fluid-structure interaction method, Acta Mech. Sin., 23 (2007), 359-367. |
| [14] | Y. Liu, S. Li, X. Sun, Numerical analysis of ossicular chain lesion of human ear, Acta Mech. Sin., 2 (2009), 241-247. |
| [15] | D. S. Arick, S. Silman, Treatment of otitis media with effusion based on politzerization with an automated device, Ear Nose Throat J., 79 (2000), 290-296. |
| [16] | J. D. Swarts, S. Foley, C. M. Alper, J. D. William, Mastoid geometry in a cross-section of humans from infancy through early adulthood with a confirmed history of otitis media, Int. J. Pediatr. Otorhinolaryngol., 76 (2012), 137-141. |
| [17] | G. Volandri, P. F. Di, P. Forte, S. Manetti, Model-oriented review and multi-body simulation of the ossicular chain of the human middle ear, Med. Eng. Phys., 34 (2012), 1339-1355. |
| [18] | S. N. Ghadiali, J. D. Swarts, W. J. Federspiel, Model-based evaluation of Eustachian tube mechanical properties using continuous pressure-flow rate data, Ann. Biomed. Eng., 30 (2002), 1064-1076. |
| [19] | F. J. Sheer, J. D. Swarts, S. N. Ghadiali, Three-dimensional finite element analysis of Eustachian tube function under normal and pathological conditions, Med. Eng. Phys., 34 (2012), 605-616. |
| [20] | M. Sudo, I. Sando, C. Suzuki, Three-dimensional reconstruction and measurement study of human Eustachian tube structures: a hypothesis of Eustachian tube function, Ann. Otol. Rhinol. Laryngol., 107 (1998), 547-554. |
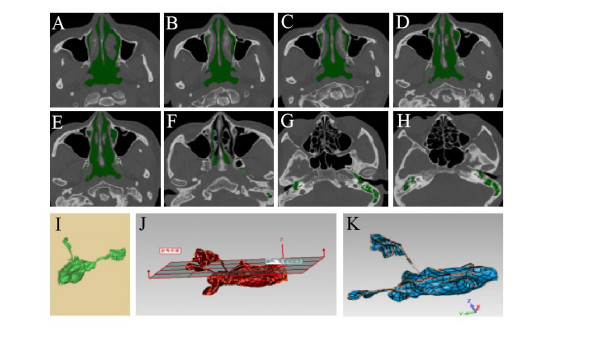
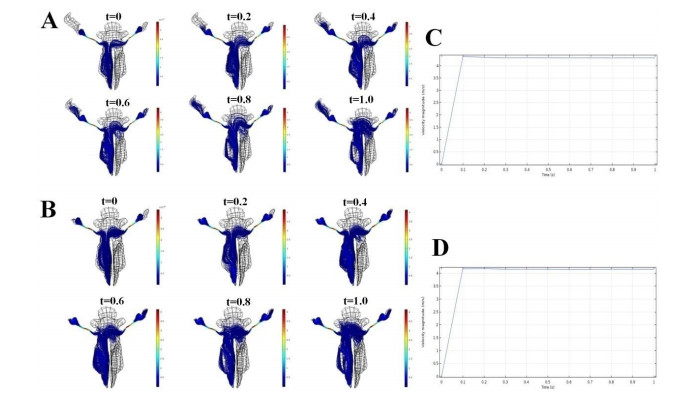
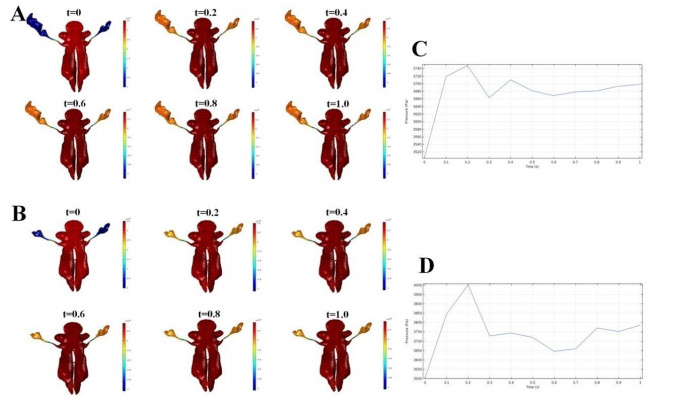
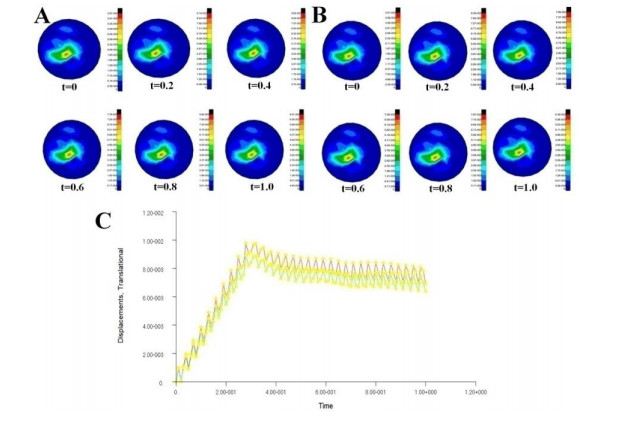
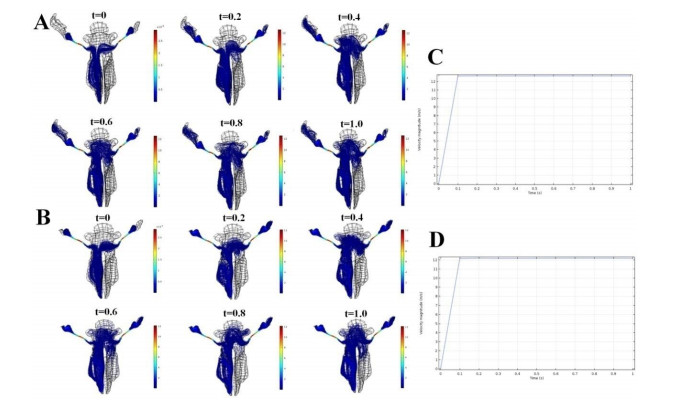
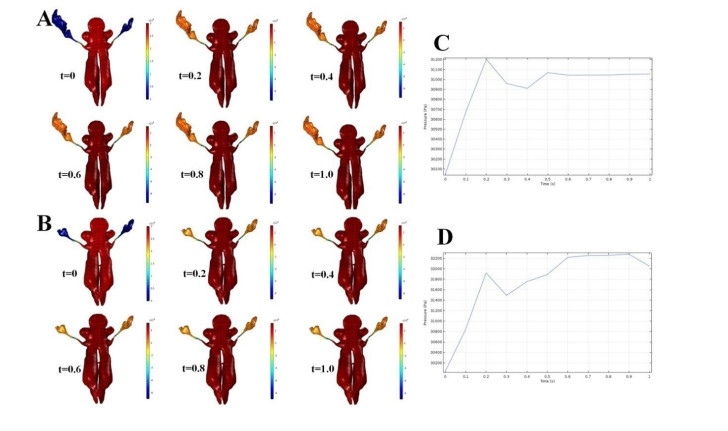
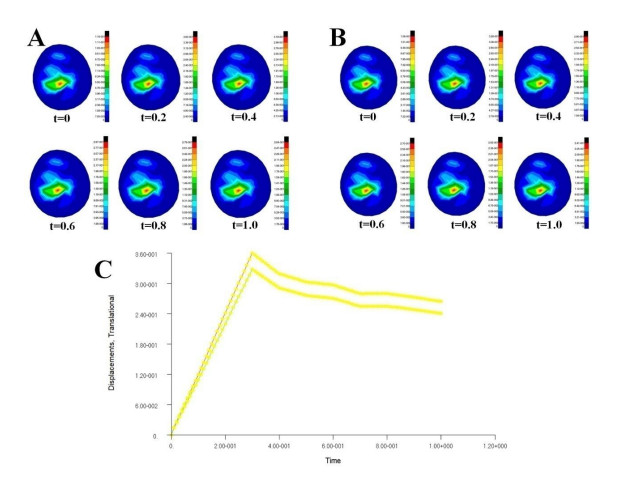
Figures(7) / Tables(3)

Xu Bie, Yuanyuan Tang, Ming Zhao, Yingxi Liu, Shen Yu, Dong Sun, Jing Liu, Ying Wang, Jianing Zhang, Xiuzhen Sun. Pilot study of pressure-flow properties in a numerical model of the middle ear[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2418-2431. doi: 10.3934/mbe.2020131







 DownLoad:
DownLoad: