Citation: Guilherme M Lopes, José F Fontanari. Influence of technological progress and renewability on the sustainability of ecosystem engineers populations[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3450-3464. doi: 10.3934/mbe.2019173

| [1] | T. R. Malthus, An essay on the theory of population, Oxford University Press, Oxford, 1798. |
| [2] | D. H. Meadows, D. L. Meadows, J. Randers, et al., The Limits to Growth; A Report for the Club of Rome's Project on the Predicament of Mankind, Universe Books, New York, 1972. |
| [3] | D. H. Meadows, J. Randers and D. L. Meadows, Limits To Growth: The 30-Year Update, Chelsea Green Publishing, Vermont, 2004. |
| [4] | J. Randers, 2052: A Global Forecast for the Next Forty Years, Chelsea Green Publishing, Vermont, 2012. |
| [5] | J. D. Murray, Mathematical Biology: I. An Introduction, 3 rd edition, Springer, New York, 2003. |
| [6] | J. A. Brander and M. S. Taylor, The Simple Economics of Easter Island: A Ricardo-Malthus Model of Renewable Resource Use, Am. Econ. Rev., 88 (1998), 119–138. |
| [7] | S. Motesharrei, J. Rivas and E. Kalnay, Human and nature dynamics (HANDY): Modeling inequality and use of resources in the collapse or sustainability of societies, Ecol. Econom., 101 (2014), 90–102. |
| [8] | B. D. Smith, The Ultimate Ecosystem Engineers,Science, 315 (2007), 1797–1798. |
| [9] | J. A. Tainter, The Collapse of Complex Societies, Cambridge University Press, Cambridge, UK, 1990. |
| [10] | J. Diamond, Collapse: How Societies Choose to Fail or Succeed, Penguin Books, New York, 2005. |
| [11] | C. G. Jones, J. H. Lawton and M. Shachak, Organisms as ecosystem engineers, Oikos, 69 (1994), 373–386. |
| [12] | W. S. C. Gurney and J. H. Lawton, The population dynamics of ecosystem engineers, Oikos, 76 (1996), 273–283. |
| [13] | J. P. Wright and C. G. Jones, The Concept of Organisms as Ecosystem Engineers Ten Years On: Progress, Limitations, and Challenges, Bioscience, 56 (2006), 203–209. |
| [14] | K. Cuddington, W. G. Wilson and A. Hastings, Ecosystem Engineers: Feedback and Population Dynamics, Am. Nat., 173 (2009), 488–498. |
| [15] | C. Franco and J. F. Fontanari, The spatial dynamics of ecosystem engineers, Math. Biosci., 292 (2017), 76–85. |
| [16] | R. J. H. Beverton and S. J. Holt, On the Dynamics of Exploited Fish Populations, Blackburn Press, Caldwell, NJ, 1993. |
| [17] | W. E. Ricker, Stock and Recruitment, J. Fish. Res. Bd. Canada, 11 (1954), 559–623. |
| [18] | J. F. Fontanari, The Collapse of Ecosystem Engineer Populations, Mathematics, 6 (2018), 9. |
| [19] | K. Kaneko, Overview of Coupled Map Lattices, Chaos, 2 (1992), 279–282. |
| [20] | M. P. Hassell, H. N. Comins and R. M. May, Spatial structure and chaos in insect population dynamics, Nature, 353 (1991), 255–258. |
| [21] | H. N. Comins, M. P. Hassell and R. M. May, The spatial dynamics of host-parasitoid systems, J. Anim. Ecol., 61 (1992), 735–748. |
| [22] | L. A. D. Rodrigues, D. C. Mistro and S. Petrovskii, Pattern Formation, Long-Term Transients, and the Turing-Hopf Bifurcation in a Space- and Time-Discrete Predator-Prey System, Bull. Math. Biol., 73 (2011), 1812–1840. |
| [23] | D. C. Mistro, L. A. D. Rodrigues and S. Petrovskii, Spatiotemporal complexity of biological invasion in a space- and time-discrete predator-prey system with the strong Allee effect, Ecol. Complex., 9 (2012), 16–32. |
| [24] | A. A. Berryman and J. A. Millstein, Are ecological systems chaotic – And if not, why not? Trends Ecol. Evol., 4 (1989), 26–28. |
| [25] | J. C. Sprott, Chaos and Time-Series Analysis, Oxford University Press, Oxford, 2003. |
| [26] | J. F. Fontanari and F. A. Rodrigues, Influence of network topology on cooperative problem-solving systems, Theory Biosci., 135 (2016), 101–110. |
| [27] | S. M. Reia and J. F. Fontanari, Effect of group organization on the performance of cooperative processes, Ecol. Complex., 30 (2017), 47–56. |
| [28] | G. Basalla, The Evolution of Technology, Cambridge University Press, Cambridge, UK, 1989. |
| [29] | F. Courchamp, J. Berec and J. Gascoigne, Allee effects in ecology and conservation, Oxford University Press, Oxford, 2008. |
| [30] | Y. A. Kuznetsov, Elements of Applied Bifurcation Theory, Springer, New York, 2004. |
| [31] | S. Strogatz, Nonlinear dynamics and chaos: with applications to physics, biology, chemistry, and engineering, Westview Press, Boulder, 2001. |
| [32] | P. Turchin, Evolution in population dynamics, Nature, 424 (2003), 257–258. |
| [33] | A. Atkisson, Believing Cassandra: How to be an Optimist in a Pessimist's World, Earthscan/Routledge, New York, 2010. |
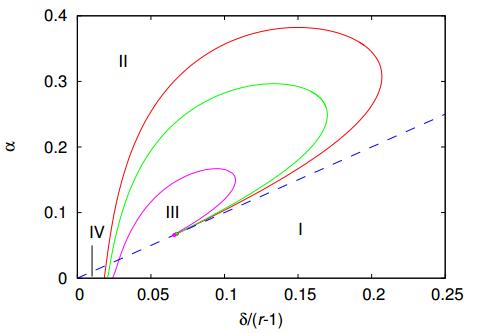
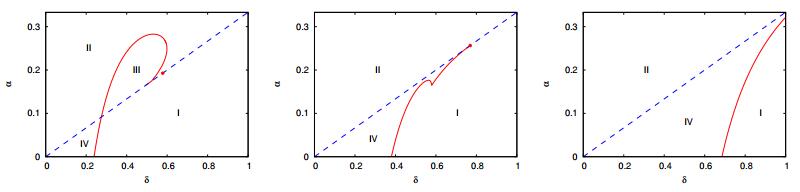
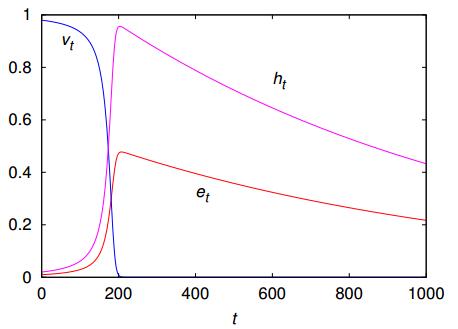
Figures(7)

Guilherme M Lopes, José F Fontanari. Influence of technological progress and renewability on the sustainability of ecosystem engineers populations[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3450-3464. doi: 10.3934/mbe.2019173







 DownLoad:
DownLoad: