Citation: Noah M. Johnson, Yuriy Y. Smolin, Chris Shindler, Daniel Hagaman, Masoud Soroush, Kenneth K. S. Lau, Hai-Feng Ji. Photochromic dye-sensitized solar cells[J]. AIMS Materials Science, 2015, 2(4): 503-509. doi: 10.3934/matersci.2015.4.503

| [1] |
Sun Z, Li H, Liu G, et al. (2014) Photochromism of New Unsymmetrical Diarylethenes Based on the Hybrid of Azaindole and Thiophene Moieties. Dye Pigment 106: 94-104. doi: 10.1016/j.dyepig.2014.03.003

|
| [2] |
Pu S, Fan C, Miao W, et al. (2008) New Photochromic Diarylethenes with a Six-Membered Aryl Unit. Tetrahedron 64: 9464-9470. doi: 10.1016/j.tet.2008.07.073
|
| [3] |
Yun C, You J, Kim J, et al. (2009) Photochromic Fluorescence Switching from Diarylethenes and Its Applications. J Photochem Photobiol C Photochem Rev 10: 111-129. doi: 10.1016/j.jphotochemrev.2009.05.002
|
| [4] |
Irie M (2010) Photochromism of Diarylethene Single Molecules and Single Crystals. Photochem Photobiol Sci 9: 1535-1542. doi: 10.1039/c0pp00251h
|
| [5] |
Bandara HMD, Burdette SC (2012) Photoisomerization in Different Classes of Azobenzene. Chem Soc Rev 41: 1809. doi: 10.1039/C1CS15179G
|
| [6] |
Samanta S, Locklin J (2008) Formation of Photochromic Spiropyran Polymer Brushes via Surface-Initiated, Ring-Opening Metathesis Polymerization: Reversible Photocontrol of Wetting Behavior and Solvent Dependent Morphology Changes. Langmuir 24: 9558-9565. doi: 10.1021/la8017387
|
| [7] |
Levy D, Einhorn S, Avnir D (1989) Applications of the Sol-Gel Process for the Preparation of Photochromic Information-Recording Materials: Synthesis, Properties, Mechanisms. J Non Cryst Solids 113: 137-145. doi: 10.1016/0022-3093(89)90004-5
|
| [8] | Keum S-R, Hur M-S, Kazmaier PM, et al. (1991) Thermo- and Photochromic Dyes: Indolino-Benzospiropyrans. Part 1. UV-VIS Spectroscopic Studies of 1,3,3-spiro(2 H-1-Benzopyran-2,2'-Indolines) and the Open-Chain Merocyanine Forms; Solvatochromism and Medium Effects on Spiro Ring Formation. Can J Chem 69: 1940-1947. |
| [9] |
Biteau J, Chaput F, Boilot J-P (1996) Photochromism of Spirooxazine-Doped Gels. J Phys Chem 100: 9024-9031. doi: 10.1021/jp953607o
|
| [10] |
Evans RA, Hanley TL, Skidmore MA, et al. (2005) The Generic Enhancement of Photochromic Dye Switching Speeds in a Rigid Polymer Matrix. Nat Mater 4: 249-253. doi: 10.1038/nmat1326
|
| [11] |
Xiaoliu L, Yongmei W, Teruo M, et al. (1999) Synthesis of New Spiropyrans and Spirooxazines Having a Heteroaromatic Pendant and Their Photochromic Behavior. Heterocycles 51: 2639-2651. doi: 10.3987/COM-99-8663
|
| [12] |
O'Regan B, Grätzel M (1991) A Low-Cost, High-Efficiency Solar Cell Based on Dye-Sensitized Colloidal TiO2 Films. Nature 353: 737-740. doi: 10.1038/353737a0
|
| [13] |
Grätzel M (2003) Dye-Sensitized Solar Cells. J Photochem Photobiol C Photochem Rev 4: 145-153. doi: 10.1016/S1389-5567(03)00026-1
|
| [14] |
Gong J, Liang J, Sumathy K (2012) Review on Dye-Sensitized Solar Cells (DSSCs): Fundamental Concepts and Novel Materials. Renew Sustain Energy Rev 16: 5848-5860. doi: 10.1016/j.rser.2012.04.044
|
| [15] |
Smolin YY, Nejati S, Bavarian M, et al. (2015) Effects of Polymer Chemistry on Polymer-Electrolyte Dye Sensitized Solar Cell Performance: A Theoretical and Experimental Investigation. J Power Sources 274: 156-164. doi: 10.1016/j.jpowsour.2014.10.028
|
| [16] |
Chen X, Tang Q, He B, et al. (2014) Platinum-Free Binary Co-Ni Alloy Counter Electrodes for Efficient Dye-Sensitized Solar Cells. Angew Chem Int Ed Engl 53: 10799-10803. doi: 10.1002/anie.201406982
|
| [17] |
Yin J-F, Velayudham M, Bhattacharya D, et al. (2012) Structure Optimization of Ruthenium Photosensitizers for Efficient Dye-Sensitized Solar Cells—A Goal toward a “bright” Future. Coord Chem Rev 256: 3008-3035. doi: 10.1016/j.ccr.2012.06.022
|
| [18] |
Hagberg DP, Marinado T, Karlsson KM, et al. (2007) Tuning the HOMO and LUMO Energy Levels of Organic Chromophores for Dye Sensitized Solar Cells. J Org Chem 72: 9550-9556. doi: 10.1021/jo701592x
|
| [19] |
Choi H, Baik C, Kang SO, et al. (2008) Highly Efficient and Thermally Stable Organic Sensitizers for Solvent-Free Dye-Sensitized Solar Cells. Angew Chemie 120: 333-336. doi: 10.1002/ange.200703852
|
| [20] |
Grätzel M (2003) Dye-Sensitized Solar Cells. J Photochem Photobiol C Photochem Rev 4: 145-153. doi: 10.1016/S1389-5567(03)00026-1
|
| [21] |
Wu W, Wang J, Zheng Z, et al. (2015) A Strategy to Design Novel Structure Photochromic Sensitizers for Dye-Sensitized Solar Cells. Sci Rep 5: 8592. doi: 10.1038/srep08592
|
| [22] |
Ma S, Ting H, Ma Y, et al. (2015) Smart Photovoltaics Based on Dye-Sensitized Solar Cells Using Photochromic Spiropyran Derivatives as Photosensitizers. AIP Adv 5: 057154. doi: 10.1063/1.4921880
|
| [23] |
Ito S, Murakami T, Comte P, et al. (2008) Fabrication of Thin Film Dye Sensitized Solar Cells with Solar to Electric Power Conversion Efficiency over 10%. Thin Solid Films 516: 4613-4619. doi: 10.1016/j.tsf.2007.05.090
|
| [24] |
Smestad GP. (1998) Demonstrating Electron Transfer and Nanotechnology: A Natural Dye-Sensitized Nanocrystalline Energy Converter. J Chem Educ 75: 752-756. doi: 10.1021/ed075p752
|
| [25] | Nazeeruddin MK, Kay A, Rodicio I, et al. (1993) Conversion of Light to Electricity by Cis-X2bis(2,2'-Bipyridyl-4,4'-dicarboxylate)ruthenium(II) Charge-Transfer Sensitizers (X = Cl-, Br-, I-, CN-, and SCN-) on Nanocrystalline Titanium Dioxide Electrodes. J Am Chem Soc 115: 6382-6390. |
| [26] |
Yu H, Zhang S, Zhao H, et al. (2009) An Efficient and Low-Cost TiO2 Compact Layer for Performance Improvement of Dye-Sensitized Solar Cells. Electrochim Acta 54: 1319-1324. doi: 10.1016/j.electacta.2008.09.025
|
| [27] |
Hore S, Vetter C, Kern R, et al. (2006) Influence of Scattering Layers on Efficiency of Dye-Sensitized Solar Cells. Sol Energy Mater Sol Cells 90: 1176-1188. doi: 10.1016/j.solmat.2005.07.002
|
| [28] |
Sommeling PM, O'Regan BC, Haswell RR, et al. (2006) Influence of a TiCl4 Post-Treatment on Nanocrystalline TiO2 Films in Dye-Sensitized Solar Cells. J Phys Chem B 110: 19191-19197. doi: 10.1021/jp061346k
|
| [29] |
Ito S, Murakami TN, Comte P, et al. (2008) Fabrication of Thin Film Dye Sensitized Solar Cells with Solar to Electric Power Conversion Efficiency over 10 Percent. Thin Solid Films 516: 4613-4619. doi: 10.1016/j.tsf.2007.05.090
|
| [30] | Tsujimoto K, Ito S, Yusa S, et al. (2011) Dye-Sensitized Solar Cells Using Surface-Stabilized Nanocrystalline-TiO2 Electrodes Coated by Epoxy Polymer. Energy Science and Technology. November 30, 2011, pp 74-78. |
| [31] |
Baumann L, Schöller K, de Courten D, et al. (2013) Development of Light-Responsive Porous Polycarbonate Membranes for Controlled Caffeine Delivery. RSC Adv 3: 23317. doi: 10.1039/c3ra44399j
|
| [32] |
Nazeeruddin MK, Zakeeruddin SM, Humphry-Baker R, et al. (1999) Acid-Base Equilibria of (2,2'-Bipyridyl-4,4'-Dicarboxylic acid)ruthenium(II) Complexes and the Effect of Protonation on Charge-Transfer Sensitization of Nanocrystalline Titania. Inorg Chem 38: 6298-6305. doi: 10.1021/ic990916a
|
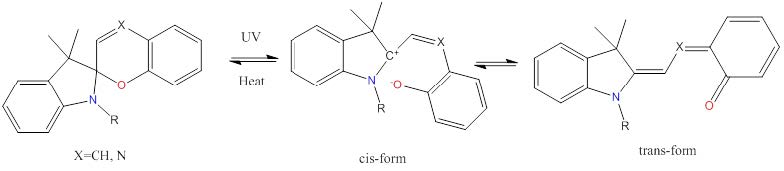
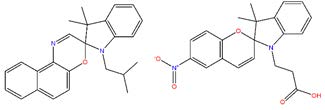

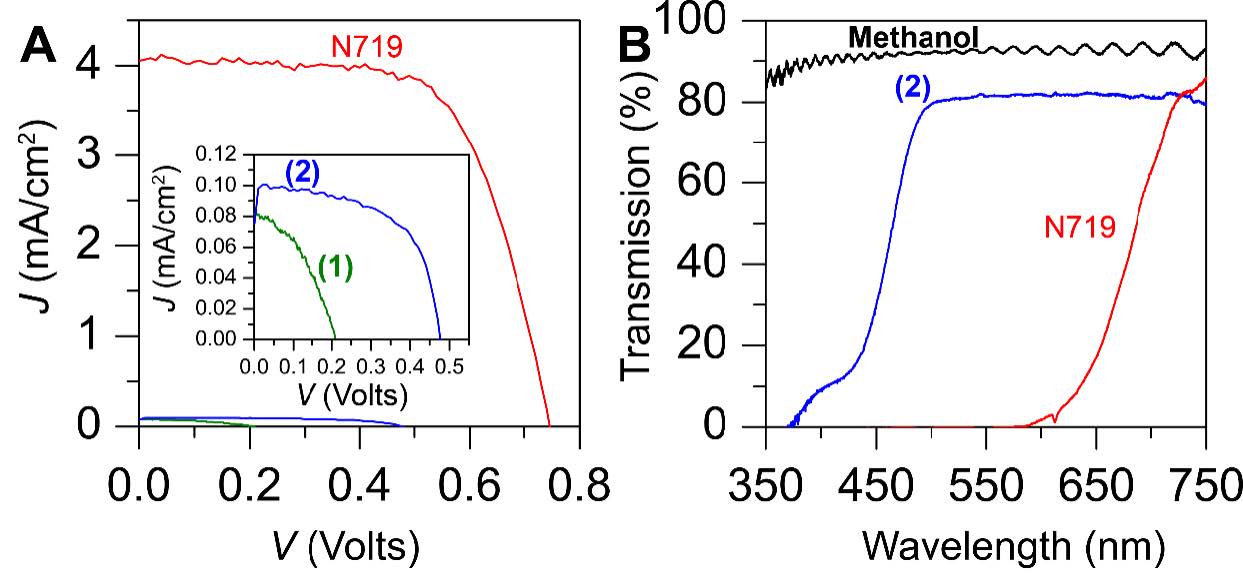
Figures(4) / Tables(1)

Noah M. Johnson, Yuriy Y. Smolin, Chris Shindler, Daniel Hagaman, Masoud Soroush, Kenneth K. S. Lau, Hai-Feng Ji. Photochromic dye-sensitized solar cells[J]. AIMS Materials Science, 2015, 2(4): 503-509. doi: 10.3934/matersci.2015.4.503







 DownLoad:
DownLoad: