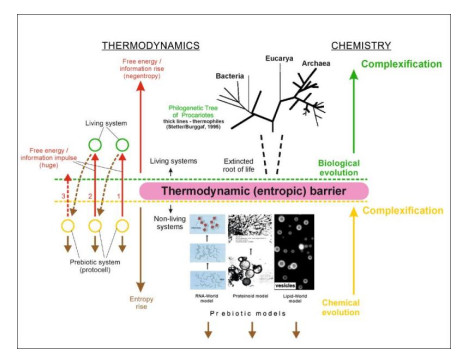
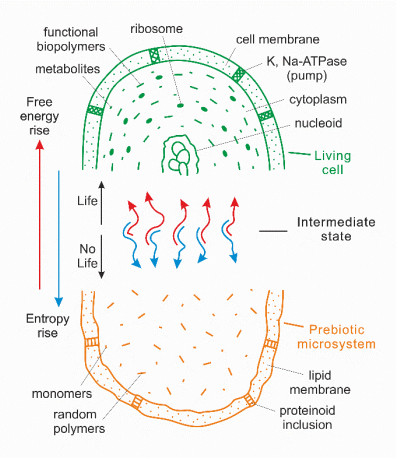
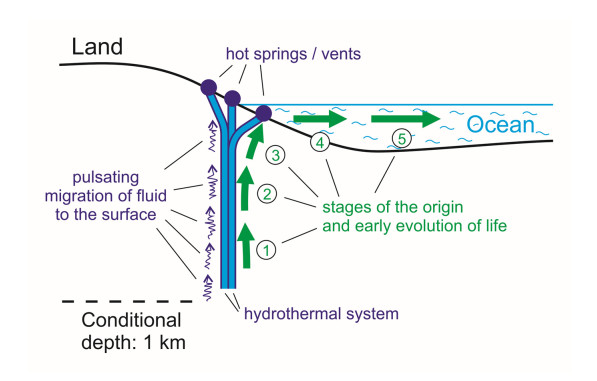
Based on the concept of thermodynamic inversion (TI), the role and place of geothermal energy (including thermal and high-frequency fluctuation energy) in the process of the emergence of the biosphere is considered, and a comparison is made with an alternative solar power source. From the standpoint of the general thermodynamics of systems, the appearance of signs of a living state in nonliving organic systems means a transition from the prevalence of the entropy contribution in the system (Sc > Fc) to the relative prevalence of the free energy contribution (Sc < Fc). Such a transition can only occur in conditions far from equilibrium in the presence of different-rank oscillations of physical and chemical parameters in the environment (temperature, pressure, chemical, and electrical potentials, etc.). Their range must include high-frequency (short-period) oscillations that bring maximum energy to the prebiotic system. Such conditions exist in hydrothermal systems during the fluid migration to the surface. Five stages of the origin of primary living cells have been identified. In the initial stages (1–3), which occurred in the subsurface areas of hydrothermal systems, self-assembly of organic microsystems and their primary evolution to the level of protolife (subcells with a primary protein-synthesizing apparatus without DNA) occurred due to geothermal energy. With the exit of subcells on the surface in geothermal regions, life (as we know it) emerged due to the involvement of solar energy into the process: a cell growth cycle appeared and formed a genetic apparatus (4–5).
Citation: Vladimir N. Kompanichenko. Geothermal energy as an initial factor in the process of life origin[J]. AIMS Geosciences, 2025, 11(2): 528-539. doi: 10.3934/geosci.2025022
Based on the concept of thermodynamic inversion (TI), the role and place of geothermal energy (including thermal and high-frequency fluctuation energy) in the process of the emergence of the biosphere is considered, and a comparison is made with an alternative solar power source. From the standpoint of the general thermodynamics of systems, the appearance of signs of a living state in nonliving organic systems means a transition from the prevalence of the entropy contribution in the system (Sc > Fc) to the relative prevalence of the free energy contribution (Sc < Fc). Such a transition can only occur in conditions far from equilibrium in the presence of different-rank oscillations of physical and chemical parameters in the environment (temperature, pressure, chemical, and electrical potentials, etc.). Their range must include high-frequency (short-period) oscillations that bring maximum energy to the prebiotic system. Such conditions exist in hydrothermal systems during the fluid migration to the surface. Five stages of the origin of primary living cells have been identified. In the initial stages (1–3), which occurred in the subsurface areas of hydrothermal systems, self-assembly of organic microsystems and their primary evolution to the level of protolife (subcells with a primary protein-synthesizing apparatus without DNA) occurred due to geothermal energy. With the exit of subcells on the surface in geothermal regions, life (as we know it) emerged due to the involvement of solar energy into the process: a cell growth cycle appeared and formed a genetic apparatus (4–5).

| [1] |
Miller SL (1953) Production of Amino Acids Under Possible Primitive Earth Conditions. Science 117: 528–529. https://doi.org/10.1126/science.117.3046.528 doi: 10.1126/science.117.3046.528

|
| [2] |
Joice GF (2002) The antiquity of RNA-world evolution. Nature 418: 214–221. https://doi.org/10.1038/418214a doi: 10.1038/418214a
|
| [3] |
Hager AJ, Pollard JD, Szostak JW (1996) Ribozymes: aiming at RNA replication and protein synthesis. Chem Biol 3: 717–725. https://doi.org/10.1016/S1074-5521(96)90246-X doi: 10.1016/S1074-5521(96)90246-X
|
| [4] |
Luisi PL (2000) The relevance of supramolecular chemistry for the origin of life. Adv Supramol Chem 6: 287–307. https://doi.org/10.1016/S1068-7459(00)80009-0 doi: 10.1016/S1068-7459(00)80009-0
|
| [5] |
Ehrenfreund P, Rasmussen S, Cleaves J, et al. (2006) Experimentally tracing the key steps in the origin of life: The aromatic world. Astrobiology 6: 490–520. https://doi.org/10.1089/ast.2006.6.490 doi: 10.1089/ast.2006.6.490
|
| [6] | Corliss JB, Baross JA, Hoffman SE (1981) An hypothesis concerning the relationship between submarine hot springs and the origin of life on the Earth. Oceanol Acta SP 4: 59–69. |
| [7] |
Holm NG, Andersson E (2005) Hydrothermal simulation experiments as a tool for studies for the origin of life on Earth and other terrestrial planets: a review. Astrobiology 5: 444–460. https://doi.org/10.1089/ast.2005.5.44 doi: 10.1089/ast.2005.5.44
|
| [8] |
Martin W, Russell JM (2007) On the origin of biochemistry at an alkaline hydrothermal vent. Philos Trans R Soc B 362: 1887–1925. https://doi.org/10.1098/rstb.2006.1881 doi: 10.1098/rstb.2006.1881
|
| [9] |
Kawamura K, Shimahashi M (2008) One-step formation of oligopeptide-like molecules from Glu and Asp in hydrothermal environments. Naturwissen 95: 449–454. https://doi.org/10.1007/s00114-008-0342-7 doi: 10.1007/s00114-008-0342-7
|
| [10] |
Cleaves HJ, Aubrey AD, Bada JL (2009) An evaluation of critical parameters for abiotic peptide synthesis in submarine hydrothermal systems. Orig Life Evol B 39: 109–126. https://doi.org/10.1007/s11084-008-9154-1 doi: 10.1007/s11084-008-9154-1
|
| [11] | Kolb VM (2016) Green Organic Chemistry and Its Interdisciplinary Applications. Boca Raton, FL: CRC Press/Taylor & Francis. |
| [12] |
Barge LM, Branscomb E, Brucato JR, et al. (2017) Thermodynamics, Disequilibrium, Evolution: Far-From-Equilibrium Geological and Chemical Considerations for Origin-Of-Life Research. Orig Life Evol Biosph 47: 39–56. https://doi.org/10.1007/s11084-016-9508-z doi: 10.1007/s11084-016-9508-z
|
| [13] |
Deamer D, Damer B, Kompanichenko V (2019) Hydrothermal Chemistry and the Origin of Cellular Life. Astrobiology 19: 1523–1537. https://doi.org/10.1089/ast.2018.1979 doi: 10.1089/ast.2018.1979
|
| [14] |
Damer B, Deamer D (2020) The Hot Spring Hypothesis for an Origin of Life. Astrobiology 20: 1–25. https://doi.org/10.1089/ast.2019.2045 doi: 10.1089/ast.2019.2045
|
| [15] |
Kompanichenko VN, Poturay VA, Shlufman KV (2015) Hydrothermal systems of Kamchatka are models of the prebiotic environment. Orig Life Evol Biosph. 45: 93–103. https://doi.org/10.1007/s11084-015-9429-2 doi: 10.1007/s11084-015-9429-2
|
| [16] |
Van Kranendonk MJ, Baumgartner R, Djokic T, et al. (2021) Elements for the Origin of Life on Land: A Deep-Time Perspective from the Pilbara Craton of Western Australia. Astrobiology 21: 39–59. https://doi.org/10.1089/ast.2019.2107 doi: 10.1089/ast.2019.2107
|
| [17] |
Deamer D (2021) Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions. Life 11: 134. https://doi.org/10.3390/life11020134 doi: 10.3390/life11020134
|
| [18] |
Russell MJ (2021) The "Water Problem"(sic), the Illusory Pond and Life's Submarine Emergence - A Review. Life 11: 429. https://doi.org/10.3390/life11050429 doi: 10.3390/life11050429
|
| [19] |
Washington J (2000) The possible role of volcanic aquifers in prebiotic genesis of organic compounds and RNA. Orig Life Evol Biosph 30: 53–79. https://doi.org/10.1023/A:1006692606492 doi: 10.1023/A:1006692606492
|
| [20] |
Kompanichenko VN (2008) Three stages of the origin-of-life process: bifurcation, stabilization and inversion. Int J Astrobiology 7: 27–46. https://doi.org/10.1017/S1473550407003953 doi: 10.1017/S1473550407003953
|
| [21] | Kompanichenko VN (2017) Thermodynamic Inversion: Origin of Living Systems. Cham (Switzerland): Springer International Publishing. |
| [22] |
Kompanichenko V (2019) The Rise of A Habitable Planet: Four Required Conditions for the Origin of Life in the Universe. Geosciences 9: 92. https://doi.org/10.3390/geosciences9020092 doi: 10.3390/geosciences9020092
|
| [23] | Kompanichenko V (2023) Abiotic-to-biotic transition of organic systems starting with thermodynamic inversion. In: Kolb V, Clark B, Guidebook for Systems Applications in Astrobiology. CRC Press, 164–176. https://doi.org/10.1201/9781003294276-8 |
| [24] | Ebeling W, Engel A, Feistel R (1990) Physik der Evolutionsprozesse. Berlin: Akademie-Verlag. |
| [25] |
Lin SK (1996) Correlation of entropy with similarity and symmetry. J Chem Inf Comp Sci 36: 367–376. https://doi.org/10.1021/ci950077k doi: 10.1021/ci950077k
|
| [26] |
Bauer E (1920) Die Definition des Lebewesens auf Grund seiner thermodynamischen Eigenschaften und die daraus folgenden biologischen Grundprinzipien. Naturwissenschaften 8: 338–340. https://doi.org/10.1007/BF02448266 doi: 10.1007/BF02448266
|
| [27] | Bauer ES (1935) Theoretical Biology. VIEM, Moscow and Leningrad. |
| [28] |
Igamberdiev AU (2024) Biological thermodynamics: Ervin Bauer and the unification of life sciences and physics. BioSystems 235: 105089. https://doi.org/10.1016/j.biosystems.2023.105089 doi: 10.1016/j.biosystems.2023.105089
|
| [29] |
Elek G, Müller M (2024) Ervin Bauer's concept of biological thermodynamics and its different evaluations. BioSystems 235: 105090. https://doi.org/10.1016/j.biosystems.2023.105090 doi: 10.1016/j.biosystems.2023.105090
|
| [30] |
Kompanichenko V, El-Registan G (2022) Advancement of the TI concept: defining the origin-of-life stages based on the succession of a bacterial cell exit from anabiosis. AIMS Geosci 8: 398–437. https://doi.org/10.3934/geosci.2022023 doi: 10.3934/geosci.2022023
|
| [31] |
Kompanichenko V (2019) Exploring the Kamchatka Geothermal Region in the Context of Life's Beginning. Life 9:41. https://doi.org/10.3390/life9020041 doi: 10.3390/life9020041
|
| [32] |
Spitzer J (2017) Emergence of Life on Earth: A Physicochemical Jigsaw Puzzle. J Mol Evol 84:1–7. https://doi.org/10.1007/s00239-016-9775-3 doi: 10.1007/s00239-016-9775-3
|
| [33] |
Spitzer J, Pielak GJ, Poolman B (2015) Emergence of Life: Physical Chemistry Changes the Paradigm. Biol Direct 10: 33. https://doi.org/10.1186/s13062-015-0060-y doi: 10.1186/s13062-015-0060-y
|
| [34] |
Woese CR (1987) Microbial evolution. Microbiol Rev 51: 221–270. https://doi.org/10.1128/mr.51.2.221-271.1987 doi: 10.1128/mr.51.2.221-271.1987
|
Figures(3) / Tables(1)

Vladimir N. Kompanichenko. Geothermal energy as an initial factor in the process of life origin[J]. AIMS Geosciences, 2025, 11(2): 528-539. doi: 10.3934/geosci.2025022







 DownLoad:
DownLoad: