Citation: Xiaojuan Wang, Jianghong Wu, Zhongren Yang, Fenglan Zhang, Hailian Sun, Xiao Qiu, Fengyan Yi, Ding Yang, Fengling Shi. Physiological responses and transcriptome analysis of the Kochia prostrata (L.) Schrad. to seedling drought stress[J]. AIMS Genetics, 2019, 6(2): 17-35. doi: 10.3934/genet.2019.2.17

| [1] |
Engelbrecht BMJ, Comita LS, Condit R, et al. (2007) Drought sensitivity shapes species distribution patterns in tropical forests. Nature 447: 80–82. doi: 10.1038/nature05747

|
| [2] | Flato G, Gregory M, Jochem A, et al. (2013) Evaluation of climate models. In: Climate Change. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, 5 Eds., Cambridge University Press, Seiten, 741–866. |
| [3] | Anderegg WRL, Kane JM, Anderegg LDL (2012) Consequences of widespread tree mortality triggered by drought and temperature stress. Nat Clim Change 3: 30–36. |
| [4] | Famiglietti JS, Rodell M (2013) Environmental science. Water in the balance. Science 340: 1300–1301. |
| [5] |
Pinheiro C, Chaves MM (2011) Photosynthesis and drought: Can we make metabolic connections from available data. J Exp Bot 62: 869–882. doi: 10.1093/jxb/erq340
|
| [6] |
Davis AM (1979) Forage quality of Prostrata Kochia compared with three browse species. Agron J 71: 822–824. doi: 10.2134/agronj1979.00021962007100050028x
|
| [7] | Wang XJ, Sun HL, Yang D, et al. (2015) Excellent feeding plants in arid region-Kochia prostrate. Grassl Prataculture 27: 10–12. |
| [8] | Alata Z, Zhao SY, Li JZ (2010) The Kochia prostrate and its cultivation techniques. Anim Husb Feed Sci 31: 268–270. |
| [9] | Yi F, Wang XJ, Qiu X, et al. (2016) Effect of Temperature, NaCI and PEG Stress on Germination and Seedling Growth of Kochia prostrate (L.) Schrad. var.horxina D.S. Wen Seed 35: 46–50. |
| [10] | Hajihashemi S, Geuns JM, Ehsanpour AA (2013) Gene transcription of steviol glycoside biosynthesis in Stevia. rebaudiana Bertoni under polyethylene glycol, paclobutrazol and gibberellic acid treatments in vitro. Acta Physiol Plant 35: 2009–2011. |
| [11] |
He Q, Zhao S, Ma Q, et al. (2014) Endogenous salicylic acid levels and signaling positively regulate Arabidopsis response to polyethylene glycol-simulated drought stress. Plant Growth Regul 33: 871–880. doi: 10.1007/s00344-014-9438-9
|
| [12] |
Abbaspour J, Ehsanpour AA (2016) Physiological targets of salicylic acid on Artemisia aucheri BOISS as a medicinaland aromatic plant grown under in vitro drought stress. Bot Stud 57: 39. doi: 10.1186/s40529-016-0154-6
|
| [13] | Wang X, Huang J (2015) Plant Physiology and Biochemistry Experiment Technology, In: Wang X, Huang J, Principles and Techniques of Plant Physiology and Biochemistry. 3 Eds., Beijing: Higher Education Press, 131–133. |
| [14] | Li HS (2000) Principles and Techniques of Plant Physiology and Biochemistry Experiment. 2 Eds., Beijing: Higher Education Press, 46–57. |
| [15] |
Young MD, Wakefield MJ, Smyth GK, et al. (2010) Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol 11: R14. doi: 10.1186/gb-2010-11-2-r14
|
| [16] | Kanehisa M, Araki M, Goto S, et al. (2008) KEGG for linking genomes to life and the environment. Nucleic Acids Res 36: D480–D484. |
| [17] |
Mao X, Cai T, Olyarchuk JG, et al. (2005) Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 21: 3787–3793. doi: 10.1093/bioinformatics/bti430
|
| [18] | Chandrasekar V, Sairam RK, Srivastava GC (2000) Physiological and biochemical responses of hexaploid and hetraploid wheat to drought tress. Agron Crop Sci 185: 219–227. |
| [19] | McMichael BL, Elmore CD (1997) Proline accumulation in water stressed cotton leaves. Crop Sci 17: 905–908. |
| [20] |
Liu FL, Jensen CR, Anderson MN, et al. (2004) Drought stress effect on carbohydrate concentration in soybean leaves andpods during early reproductive development: Its implication in altering pod set. Field Crops Res 86: 1–13. doi: 10.1016/S0378-4290(03)00165-5
|
| [21] |
Ohashi Y, Nakayama N, Saneoka H, et al. (2006) Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biologia Plantarum 50: 138–141. doi: 10.1007/s10535-005-0089-3
|
| [22] |
Taylor SH, Franks PJ, Hulme SP, et al. (2012) Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses. New Phytologist 193: 387–396. doi: 10.1111/j.1469-8137.2011.03935.x
|
| [23] |
Franks PJ, Beerling DJ (2009) Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proceedings of the National Academy of Sciences of the United States of America 106: 10343–10347. doi: 10.1073/pnas.0904209106
|
| [24] |
Liu W, Ji SX, Fang XL, et al. (2013) Protein kinase LTRPK1 influences cold adaptation and microtubule stability in rice. Plant Growth Regul 32: 483–490. doi: 10.1007/s00344-012-9314-4
|
| [25] | Park SY, Fung P, Nishimura N, et al. (2009) Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 324: 1068–1071. |
| [26] |
Zhang X, Miao YC, An GY, et al. (2001) Kannels inhibited by hydrogen peroxide mediate abscisic acid signaling in guard cells. Cell Res 11: 195–202 doi: 10.1038/sj.cr.7290086
|
| [27] |
Rodriguez MC, Petersen M, Mundy J (2010) Mitogen-activated protein kinase signaling in plants. Annu. Rev Plant Biol 61: 621–649. doi: 10.1146/annurev-arplant-042809-112252
|
| [28] |
Bowler C, Slooten L, Vandenbranden S, et al. (1991) Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J 10: 1723–1732. doi: 10.1002/j.1460-2075.1991.tb07696.x
|
| [29] |
Jin H, Huang F, Cheng H, et al. (2013) Overexpression of the GmNAC2 gene, an NAC transcription factor, reduces abiotic stress tolerance in tobacco. Plant Mol Biol Rep 31: 435–442. doi: 10.1007/s11105-012-0514-7
|
| [30] |
Sun J, Hu W, Zhou R, et al. (2015) The Brachypodium distachyon BdWRKY36 gene confers tolerance to drought stress in transgenic tobacco plants. Plant Cell Rep 34: 23–35. doi: 10.1007/s00299-014-1684-6
|
| [31] |
Shafi A, Chauhan R, Gill T, et al. (2015) Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol Biol 87: 615–631. doi: 10.1007/s11103-015-0301-6
|
| [32] |
Xu K, Chen S, Li T, et al. (2015) OsGRAS23, a rice GRAS transcription factor gene, is involved in drought stress response through regulating expression of stress-responsive genes. BMC Plant Biol 15: 141. doi: 10.1186/s12870-015-0532-3
|
| [33] | Jin H, Huang F, Cheng H, et al. (2015) Overex-pression of the GmNAC2 gene, an NAC transcription factor, reduces abiotic stress tolerance in tobacco. Plant Mol Biol Rep 31: 435–442. |
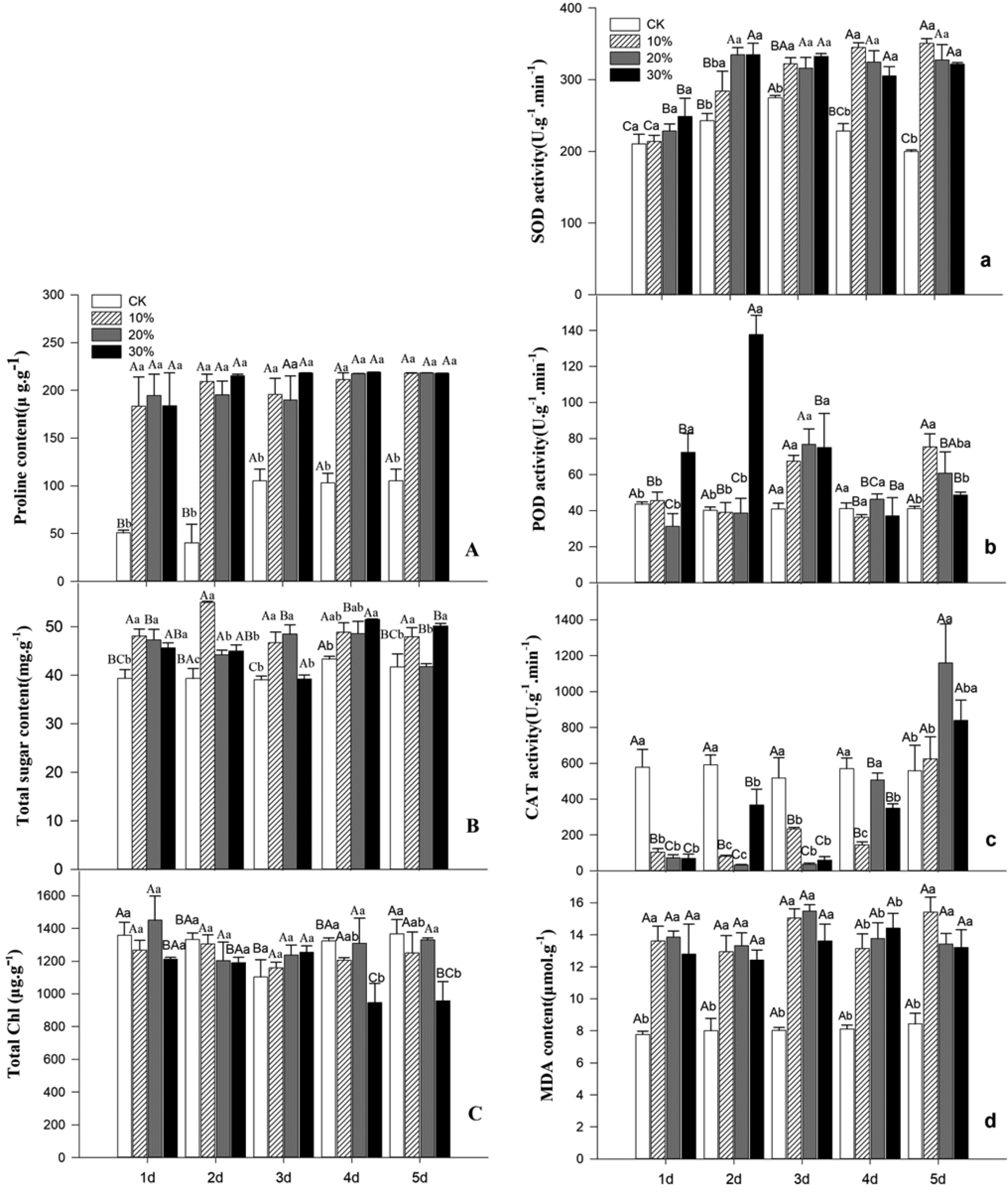
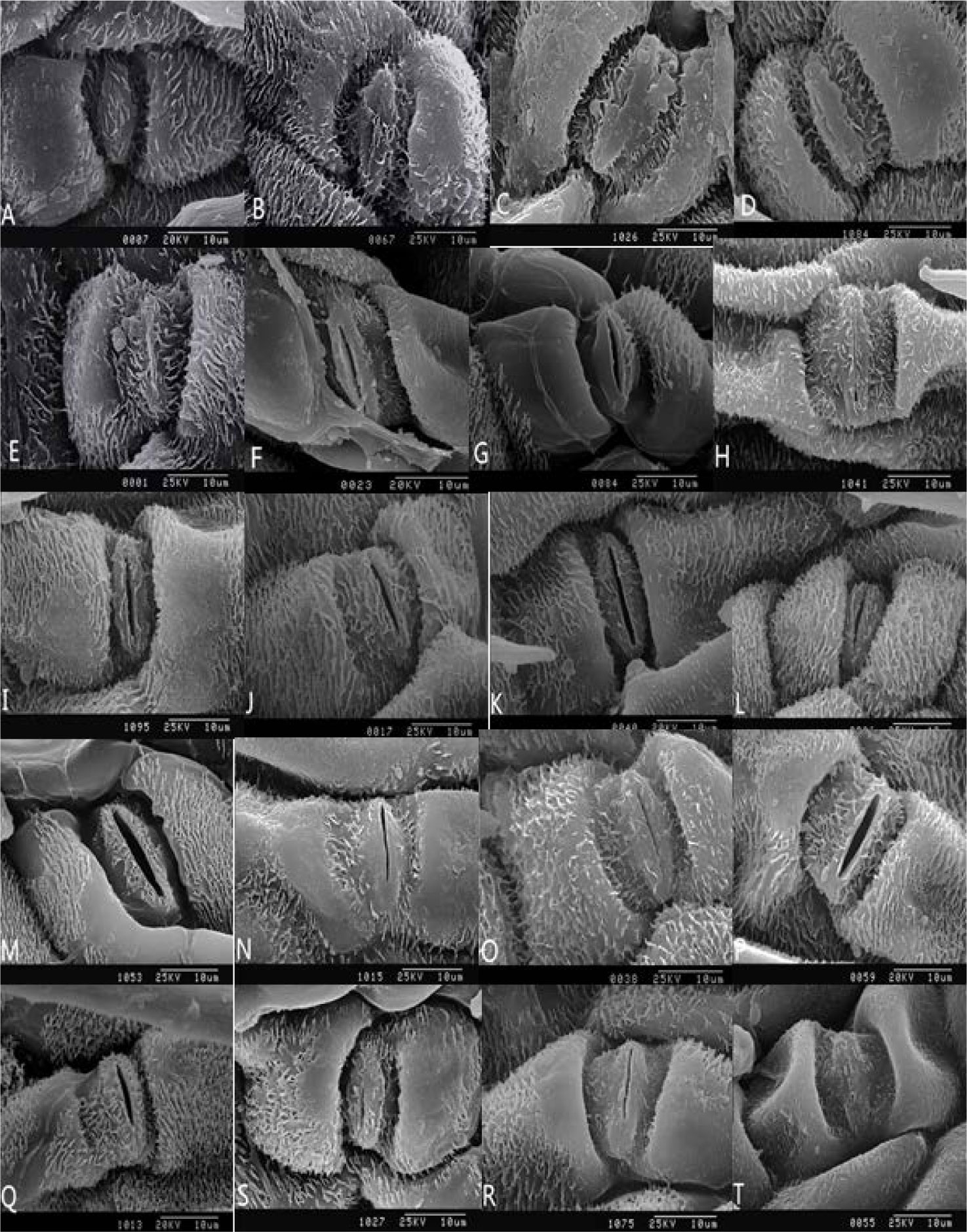
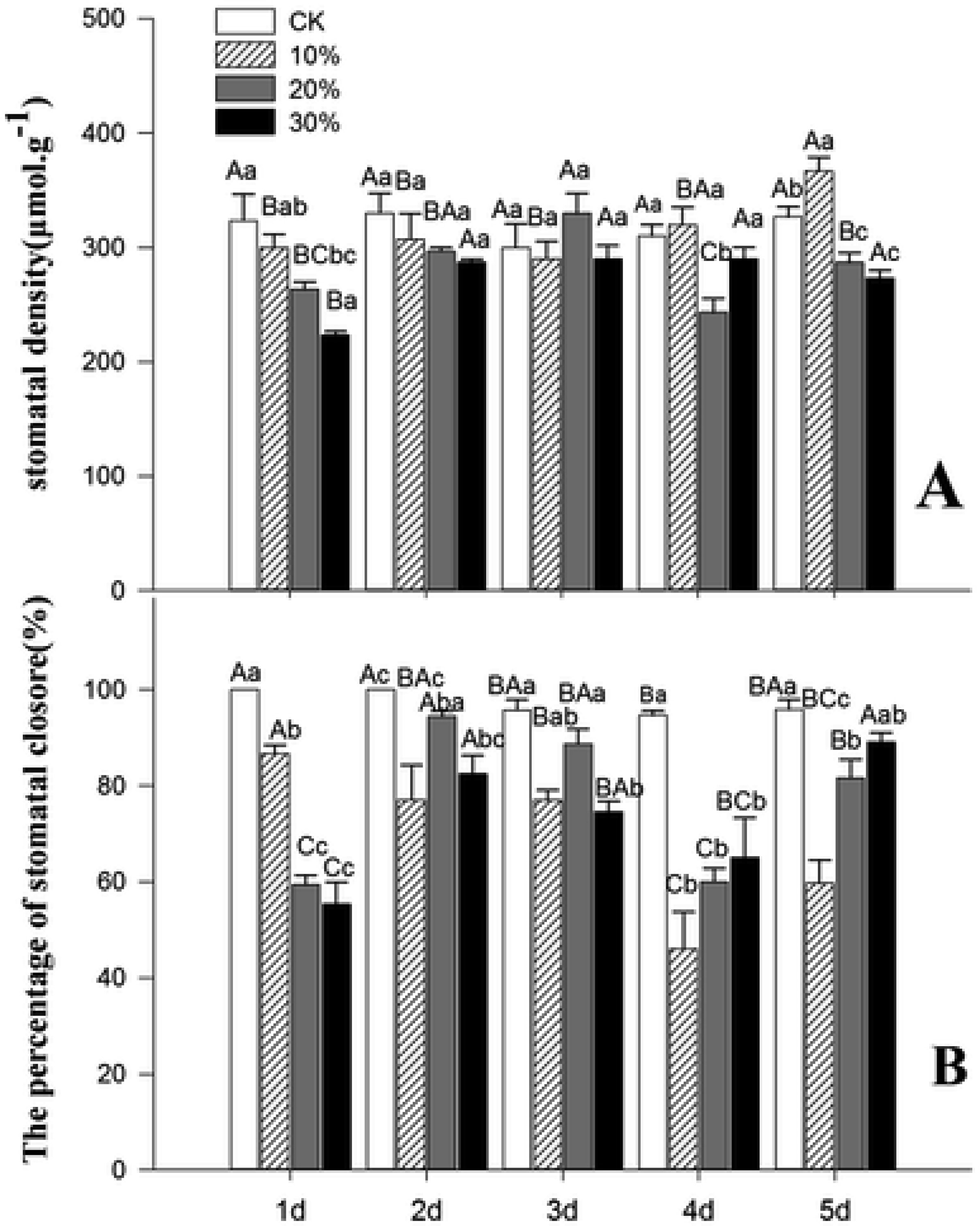
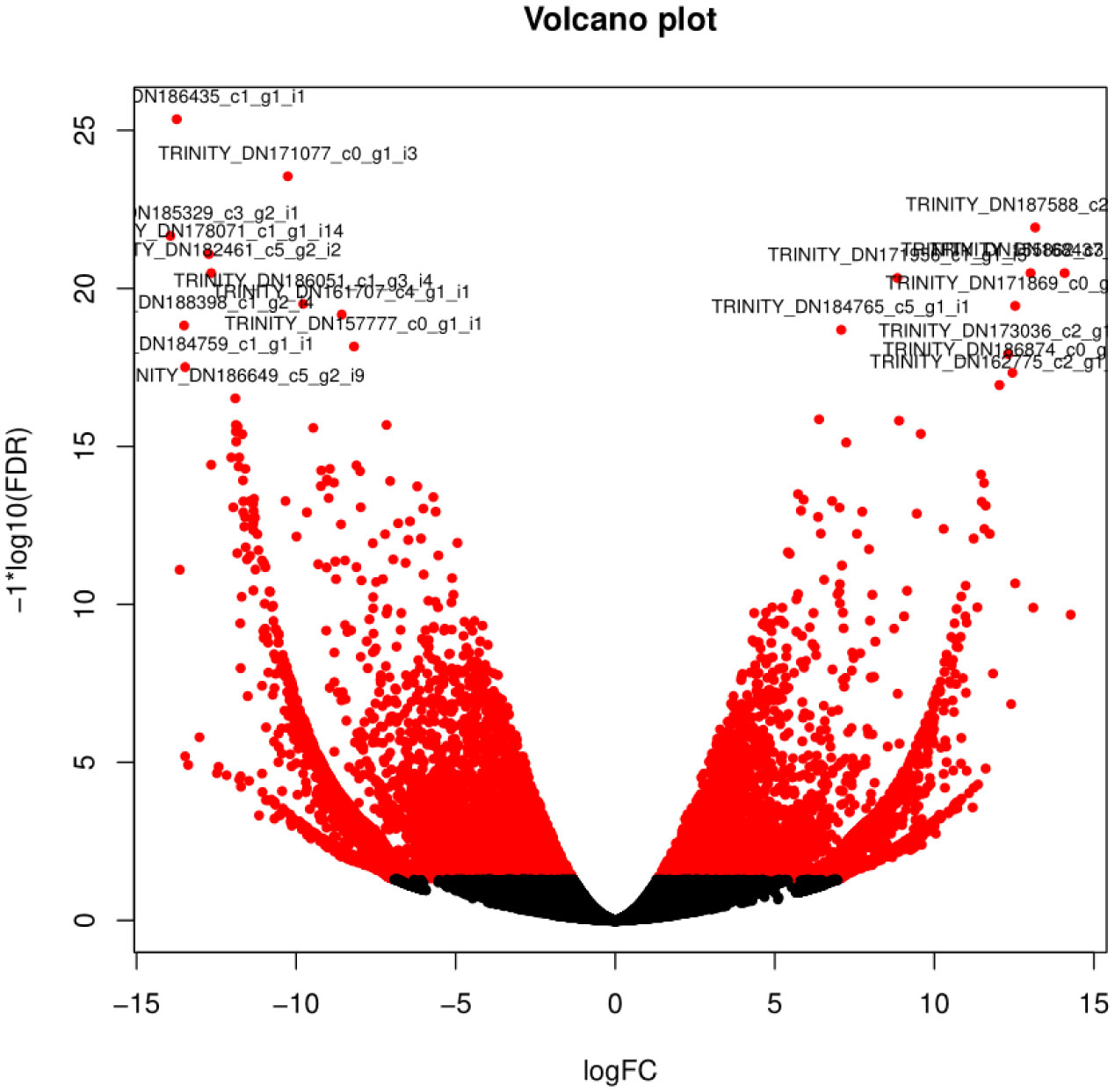
Figures(7) / Tables(2)

Xiaojuan Wang, Jianghong Wu, Zhongren Yang, Fenglan Zhang, Hailian Sun, Xiao Qiu, Fengyan Yi, Ding Yang, Fengling Shi. Physiological responses and transcriptome analysis of the Kochia prostrata (L.) Schrad. to seedling drought stress[J]. AIMS Genetics, 2019, 6(2): 17-35. doi: 10.3934/genet.2019.2.17







 DownLoad:
DownLoad: