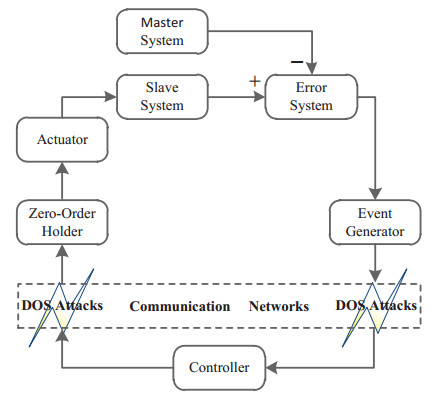
This paper studied the event-triggered synchronization problem for time-delay neural networks under DoS attacks. A novel event-triggered scheme based on switching between periodic sampling and a continuous event-triggered scheme was proposed, which not only cuts down the number of data transmissions but also offsets cyberattacks. By choosing a suitable piecewise Lyapunov-Krasovskii functional and using several free-weighting matrices, sufficient conditions were established to ensure the exponential stability of the synchronization error system in the occurrence of DoS attacks. Furthermore, a co-design method was provided to acquire the desired non-fragile output-feedback control gain and event-triggering parameter. Finally, a numerical example was given to illustrate the usefulness of the proposed approach.
Citation: Yawei Liu, Guangyin Cui, Chen Gao. Event-triggered synchronization control for neural networks against DoS attacks[J]. Electronic Research Archive, 2025, 33(1): 121-141. doi: 10.3934/era.2025007
This paper studied the event-triggered synchronization problem for time-delay neural networks under DoS attacks. A novel event-triggered scheme based on switching between periodic sampling and a continuous event-triggered scheme was proposed, which not only cuts down the number of data transmissions but also offsets cyberattacks. By choosing a suitable piecewise Lyapunov-Krasovskii functional and using several free-weighting matrices, sufficient conditions were established to ensure the exponential stability of the synchronization error system in the occurrence of DoS attacks. Furthermore, a co-design method was provided to acquire the desired non-fragile output-feedback control gain and event-triggering parameter. Finally, a numerical example was given to illustrate the usefulness of the proposed approach.

| [1] |
S. Lakshmanan, M. Prakash, C. P. Lim, R. Rakkiyappan, P. Balasubramaniam, S. Nahavandi, Synchronization of an inertial neural network with time-varying delays and its application to secure communication, IEEE Trans. Neural Networks Learn. Syst., 29 (2018), 195–207. https://doi.org/10.1109/TNNLS.2016.2619345 doi: 10.1109/TNNLS.2016.2619345

|
| [2] |
H. Huang, G. Feng, J. Cao, An LMI approach to delay-dependent state estimation for delayed neural networks, Neurocomputing, 71 (2008), 2857–2867. https://doi.org/10.1016/j.neucom.2007.08.008 doi: 10.1016/j.neucom.2007.08.008
|
| [3] |
Y. Liu, Z. Wang, X. Liu, Global exponential stability of generalized recurrent neural networks with discrete and distributed delays, Neural Networks, 19 (2006), 667–675. https://doi.org/10.1016/j.neunet.2005.03.015 doi: 10.1016/j.neunet.2005.03.015
|
| [4] |
Y. Zhao, H. Wu, Fixed/Prescribed stability criterions of stochastic system with time-delay, AIMS Math., 9 (2024), 14425–14453. https://doi.org/10.3934/math.2024701 doi: 10.3934/math.2024701
|
| [5] | Z. Yan, X. Huang, Y. Fan, J. Xia, H. Shen, Threshold-function-dependent quasi-synchronization of delayed memristive neural networks via hybrid event-triggered control, IEEE Trans. Syst. Man Cybern.: Syst., 51, (2021), 6712–6722. https://doi.org/10.1109/TSMC.2020.2964605 |
| [6] |
C. Yi, J. Cai, R. Guo, Synchronization of a class of nonlinear multiple neural networks with delays via a dynamic event-triggered impulsive control strategy, Electron. Res. Arch., 32 (2024), 4581–4603. https://doi.org/10.3934/era.2024208 doi: 10.3934/era.2024208
|
| [7] |
X. Hou, H. Wu, J. Cao, Observer-based prescribed-time synchronization and topology identification for complex networks of piecewise-smooth systems with hybrid impulses, Comp. Appl. Math., 43 (2024), 1–19. https://doi.org/10.1007/s40314-024-02701-x doi: 10.1007/s40314-024-02701-x
|
| [8] |
W. Zhao, K. Li, Y. Shi, Exponential synchronization of neural networks with mixed delays under impulsive control, Electron. Res. Arch., 32 (2023), 5286–5305. https://doi.org/10.3934/era.2024244 doi: 10.3934/era.2024244
|
| [9] |
H. Ren, Z. Peng, Y. Gu, Fixed-time synchronization of stochastic memristor-based neural networks with adaptive control, Neural Networks, 130 (2020), 165–175. https://doi.org/10.1016/j.neunet.2020.07.002 doi: 10.1016/j.neunet.2020.07.002
|
| [10] |
L. Wang, H. He, Z. Zeng, Global synchronization of fuzzy memristive neural networks with discrete and distributed delays, IEEE Trans. Fuzzy Syst., 28 (2020), 2022–2034. https://doi.org/10.1109/TFUZZ.2019.2930032 doi: 10.1109/TFUZZ.2019.2930032
|
| [11] |
X. Yang, Y. Liu, J. Cao, L. Rutkowski, Synchronization of coupled time-delay neural networks with mode-dependent average dwell time switching, IEEE Trans. Neural Networks Learn. Syst., 31 (2020), 5483–5496. https://doi.org/10.1109/TNNLS.2020.2968342 doi: 10.1109/TNNLS.2020.2968342
|
| [12] |
J. Zhou, J. Dong, X. Su, C. K. Ahn, Input-to-state stabilization for Markov jump systems with dynamic quantization and multimode injection attacks, IEEE Trans. Syst. Man Cybern.: Syst., 54 (2024), 2517–2529. https://doi.org/10.1109/TSMC.2023.3344869 doi: 10.1109/TSMC.2023.3344869
|
| [13] |
G. Ran, C. Li, R. Sakthivel, C. Han, B. Wang, J. Liu, Adaptive event-triggered asynchronous control for interval type-2 fuzzy Markov jump systems with cyberattacks, IEEE Trans. Control Network Syst., 9 (2022), 88–99. https://doi.org/10.1109/TCNS.2022.3141025 doi: 10.1109/TCNS.2022.3141025
|
| [14] |
M. Zhu, S. Martinez, On the performance analysis of resilient networked control systems under replay attacks, IEEE Trans. Autom. Control, 59 (2014), 804–808. https://doi.org/10.1109/TAC.2013.2279896 doi: 10.1109/TAC.2013.2279896
|
| [15] |
J. Zhou, D. Xu, W. Tai, C. K. Ahn Switched event-triggered $H _\infty$ security control for networked systems vulnerable to aperiodic DoS attacks, IEEE Trans. Network Sci. Eng., 10 (2023), 2109–2123. https://doi.org/10.1109/TNSE.2023.3243095 doi: 10.1109/TNSE.2023.3243095
|
| [16] |
S. Hu, D. Yue, X. Xie, X. Chen, X. Yin, Resilient event-triggered controller synthesis of networked control systems under periodic DoS jamming attacks, IEEE Trans. Cybern., 49 (2019), 4271–4281. https://doi.org/10.1109/TCYB.2018.2861834 doi: 10.1109/TCYB.2018.2861834
|
| [17] |
S. Hu, D. Yue, Q. L. Han, X. Xie, X. Chen, C. Dou, Observer-based event-triggered control for networked linear systems subject to denial-of-service attacks, IEEE Trans. Cybern., 50 (2020), 1952–1964. https://doi.org/10.1109/TCYB.2019.2903817 doi: 10.1109/TCYB.2019.2903817
|
| [18] |
D. Liu, D. Ye, Cluster synchronization of complex networks under denial-of-service attacks with distributed adaptive strategies, IEEE Trans. Control Network Syst., 9 (2022), 334–343. https://doi.org/10.1109/TCNS.2021.3102012 doi: 10.1109/TCNS.2021.3102012
|
| [19] |
J. Liu, W. Suo, L. Zha, E. Tian, X. Xie, Security distributed state estimation for nonlinear networked systems against DoS attacks, Int. J. Robust Nonlinear Control, 30 (2020), 1156–1180. https://doi.org/10.1002/rnc.4815 doi: 10.1002/rnc.4815
|
| [20] |
S. Hu, D. Yue, X. Chen, Z. Cheng, X. Xie, Resilient $H_\infty$ filtering for event-triggered networked systems under nonperiodic DoS jamming attacks, IEEE Trans. Syst. Man Cybern.: Syst., 51 (2021), 1392–1403. https://doi.org/10.1109/TSMC.2019.2896249 doi: 10.1109/TSMC.2019.2896249
|
| [21] |
X. Chen, P. Yuan, Event-triggered generalized dissipative filtering for delayed neural networks under aperiodic DoS jamming attacks, Neurocomputing, 400 (2020), 467–479. https://doi.org/10.1016/j.neucom.2019.03.088 doi: 10.1016/j.neucom.2019.03.088
|
| [22] |
X. Chen, Y. Wang, S. Hu, Event-triggered quantized $H_\infty$ control for networked control systems in the presence of denial-of-service jamming attacks, Nonlinear Anal. Hybrid Syst., 33 (2019), 265–281. https://doi.org/10.1016/j.nahs.2019.03.005 doi: 10.1016/j.nahs.2019.03.005
|
| [23] |
N. Zhao, P. Shi, W. Xing, J. Chambers, Observer-based event-triggered approach for stochastic networked control systems under denial of service attacks, IEEE Trans. Control Network Syst., 8 (2021), 158–167. https://doi.org/10.1109/TCNS.2020.3035760 doi: 10.1109/TCNS.2020.3035760
|
| [24] |
E. Fridman, A refined input delay approach to sampled-data control, Automatica, 46 (2010), 421–427. https://doi.org/10.1016/j.automatica.2009.11.017 doi: 10.1016/j.automatica.2009.11.017
|
| [25] |
Y. Yao, J. Tan, J. Wu, X. Zhang, A unified fuzzy control approach for stochastic high-order nonlinear systems with or without state constraints, IEEE Trans. Fuzzy Syst., 30 (2022), 4530–4540. https://doi.org/10.1109/TFUZZ.2022.3155297 doi: 10.1109/TFUZZ.2022.3155297
|
| [26] |
X. Zhao, H. Wu, J. Cao, L. Wang, Prescribed-time synchronization for complex dynamic networks of piecewise smooth systems: a hybrid event-triggering control approach, Qual. Theory Dyn. Syst., 24 (2025), 11. https://doi.org/10.1007/s12346-024-01166-x doi: 10.1007/s12346-024-01166-x
|
| [27] |
W. Wu, L. He, J. Zhou, Z. Xuan, S. Arik, Disturbance-term-based switching event-triggered synchronization control of chaotic Lurie systems subject to a joint performance guarantee, Commun. Nonlinear Sci. Numer. Simul., 115 (2022), 106774. https://doi.org/10.1016/j.cnsns.2022.106774 doi: 10.1016/j.cnsns.2022.106774
|
| [28] |
Y. Cao, S. Wang, Z. Guo, T. Huang, S. Wen, Synchronization of memristive neural networks with leakage delay and parameters mismatch via event-triggered control, Neural Network, 119 (2019), 178–189. https://doi.org/10.1016/j.neunet.2019.08.011 doi: 10.1016/j.neunet.2019.08.011
|
| [29] |
Y. Tan, Y. Liu, B. Niu, S. Fei, Event-triggered synchronization control for T–S fuzzy neural networked systems with time delay, J. Franklin Inst., 357 (2020), 5934–5953. https://doi.org/10.1016/j.jfranklin.2020.03.024 doi: 10.1016/j.jfranklin.2020.03.024
|
| [30] | A. Selivanov, E. Fridman, A switching approach to event-triggered control, in 2015 54th IEEE Conference on Decision and Control (CDC), Osaka, Japan, (2015), 5468–5473. https://doi.org/10.1109/CDC.2015.7403076 |
| [31] |
P. Zeng, F. Deng, X. Liu, X. Gao, Event-triggered $H_\infty$ control for network-based uncertain Markov jump systems under dos attacks, J. Franklin Inst., 358 (2021), 2895–2914. https://doi.org/10.1016/j.jfranklin.2021.01.026 doi: 10.1016/j.jfranklin.2021.01.026
|
| [32] |
Y. Yao, J. Tan, J. Wu, X. Zhang, Event-triggered fixed-time adaptive neural dynamic surface control for stochastic non-triangular structure nonlinear systems, Inf. Sci., 569 (2021), 527–543. https://doi.org/10.1016/j.ins.2021.05.028 doi: 10.1016/j.ins.2021.05.028
|
| [33] |
Y. Yao, J. Tan, J. Wu, X. Zhang, Event-triggered fixed-time adaptive fuzzy control for state-constrained stochastic nonlinear systems without feasibility conditions, Nonlinear Dyn., 105 (2021), 403–416. https://doi.org/10.21203/rs.3.rs-322781/v1 doi: 10.21203/rs.3.rs-322781/v1
|
| [34] |
F. Yang, H. Dong, Z. Wang, W. Ren, F. E. Alsaadi, A new approach to non-fragile state estimation for continuous neural networks with time-delays, Neurocomputing, 197 (2016), 205–211. https://doi.org/10.1016/j.neucom.2016.02.062 doi: 10.1016/j.neucom.2016.02.062
|
| [35] |
H. B. Zeng, Y. He, M. Wu, J. She, Free-matrix-based integral inequality for stability analysis of systems with time-varying delay, IEEE Trans. Autom. Control, 60 (2015), 2768–2772. https://doi.org/10.1109/TAC.2015.2404271 doi: 10.1109/TAC.2015.2404271
|
| [36] |
J. Zhou, J. H. Park, Q. Ma, Non-fragile observer-based $H_\infty$ control for stochastic time-delay systems, Appl. Math. Comput., 291 (2016), 69–83. https://doi.org/10.1016/j.amc.2016.06.024 doi: 10.1016/j.amc.2016.06.024
|
| [37] |
W. Tai, X. Li. J. Zhou, S. Arik, Asynchronous dissipative stabilization for stochastic Markov-switching neural networks with completely-and incompletely-known transition rates, Neural Network, 161 (2023), 55–64. https://doi.org/10.1016/j.neunet.2023.01.039 doi: 10.1016/j.neunet.2023.01.039
|
| [38] |
J. Sun, G. P. Liu, J. Chen, D. Rees, Improved delay-range-dependent stability criteria for linear systems with time-varying delays, Automatica, 46 (2010), 466–470. https://doi.org/10.1016/j.automatica.2009.11.002 doi: 10.1016/j.automatica.2009.11.002
|
| [39] |
Z. Fei, C. Guan, H. Gao, Exponential synchronization of networked chaotic delayed neural network by a hybrid event trigger scheme, IEEE Trans. Neural Network Learn. Syst., 29 (2017), 2558–2567. https://doi.org/10.1109/TNNLS.2017.2700321 doi: 10.1109/TNNLS.2017.2700321
|
| [40] |
W. Li, H. Du, D. Ning, W. Li, S. Sun, J. Wei, Event-triggered $H_\infty$ control for active seat suspension systems based on relaxed conditions for stability, Mech. Syst. Signal Process., 149 (2024), 107210. https://doi.org/10.1016/j.ymssp.2020.107210 doi: 10.1016/j.ymssp.2020.107210
|
| [41] |
Y. Zhu, Y. Yao, Y. Kang, Y. Zhao, J. Tan, L. Gu, et al., Event-based enhancing prescribed performance control for stochastic non-triangular structure nonlinear systems: a MTBFs-based approach, Nonlinear Dyn., 113 (2025), 533–545. https://doi.org/10.1007/s11071-024-10242-5 doi: 10.1007/s11071-024-10242-5
|
| [42] |
Y. Yao, Y. Kang, Y. Zhao, P. Li, J. Tan, Prescribed-time output feedback control for cyber-physical systems under output constraints and malicious attacks, IEEE Trans. Cybern., 54 (2024), 6518–6530. https://doi.org/10.1109/TCYB.2024.3418384 doi: 10.1109/TCYB.2024.3418384
|
| [43] |
X. Hou, H. Wu, J. Cao, Practical finite-time synchronization for Lur'e systems with performance constraint and actuator faults: A memory-based quantized dynamic event-triggered control strategy, Appl. Math. Comput., 487 (2025), 129108. https://doi.org/10.1016/j.amc.2024.129108 doi: 10.1016/j.amc.2024.129108
|




Figures(5) / Tables(1)
Yawei Liu, Guangyin Cui, Chen Gao. Event-triggered synchronization control for neural networks against DoS attacks[J]. Electronic Research Archive, 2025, 33(1): 121-141. doi: 10.3934/era.2025007







 DownLoad:
DownLoad: