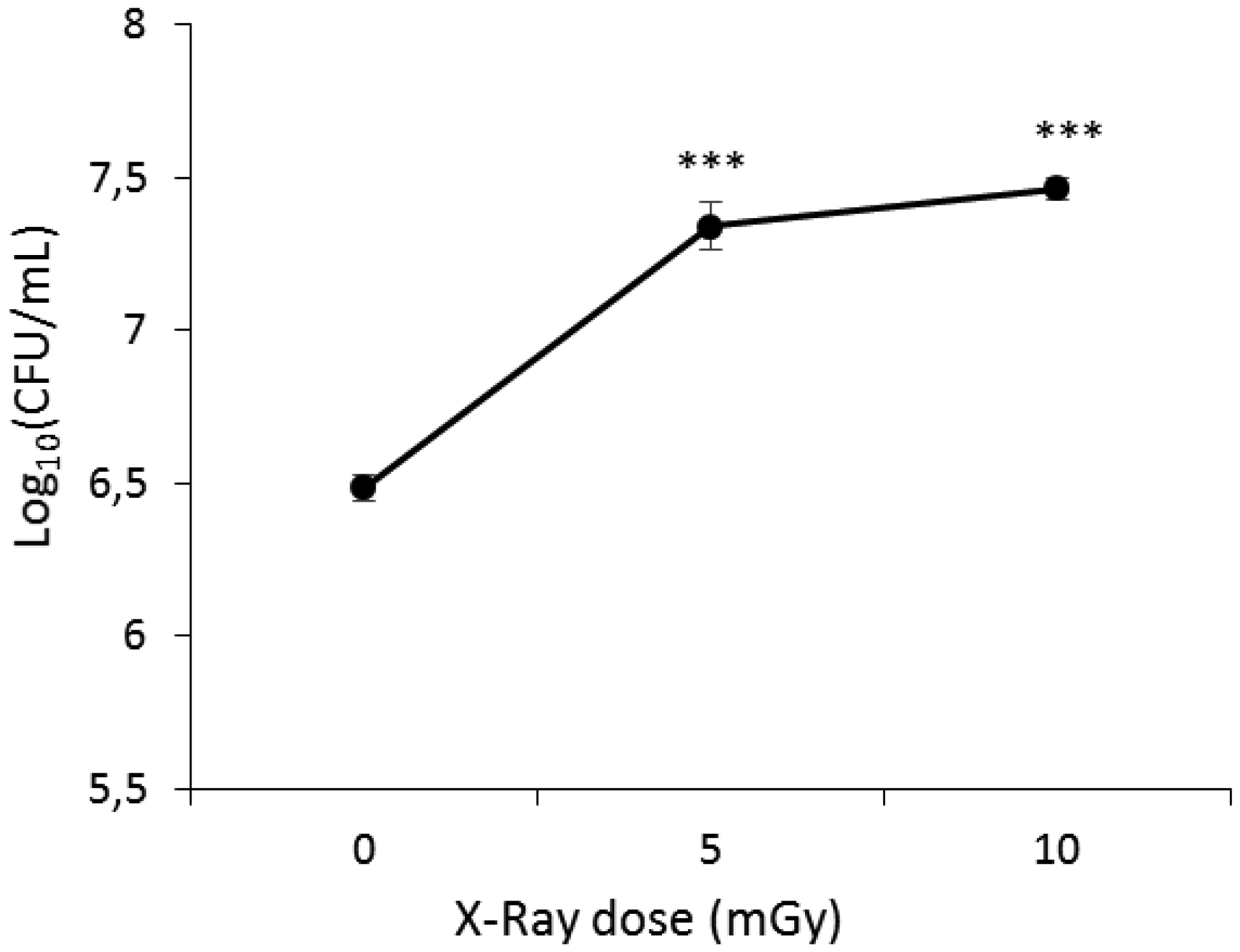
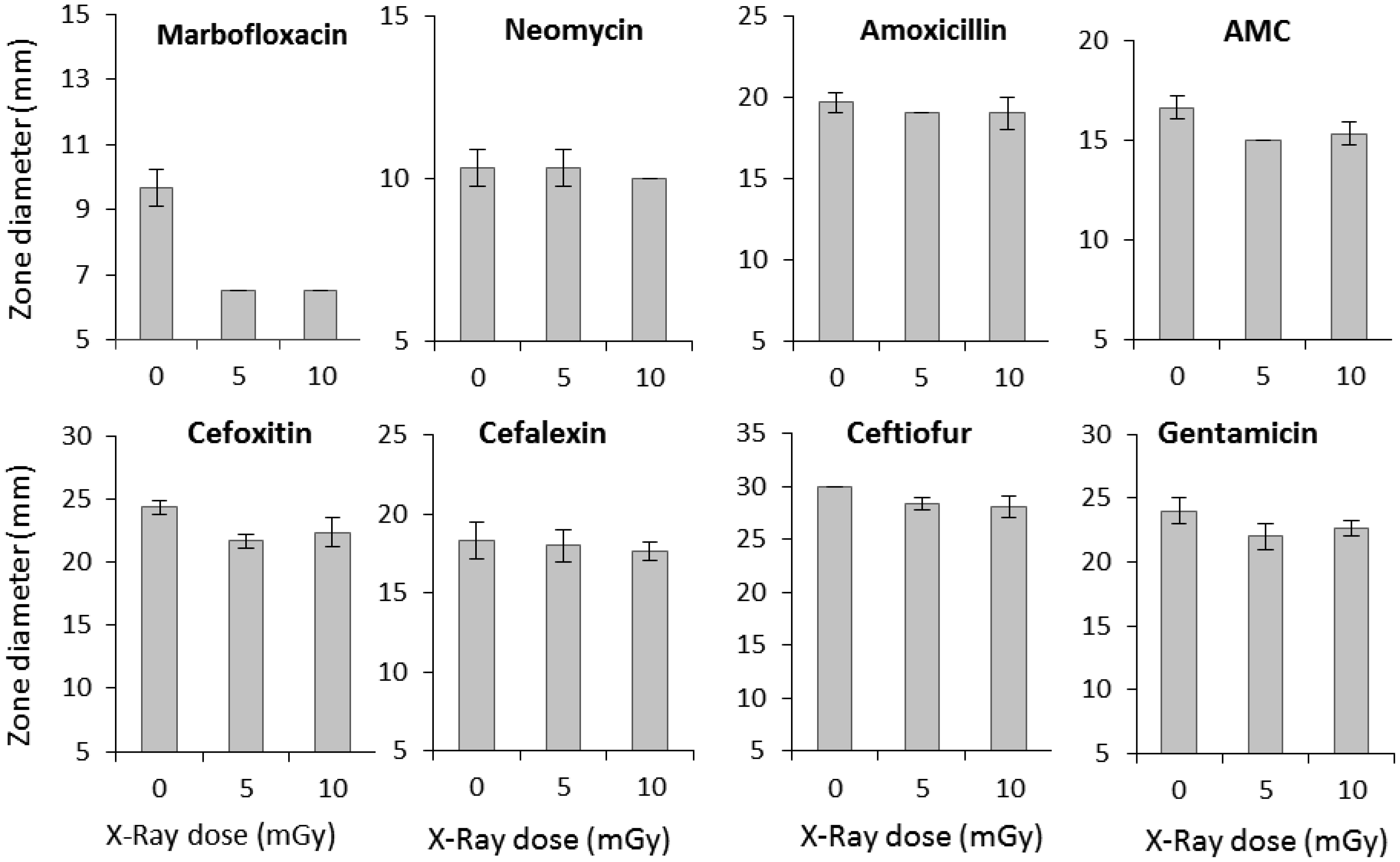
Low-dose ionizing radiation can trigger a phenomenon known as hormesis in microorganisms, in which exposure to mild stressors like radiation results in beneficial adaptive responses. This study investigated the impact of low-dose X-rays on Escherichia coli's viability and their potential influence on antibiotic susceptibility. The irradiated samples displayed increased bacterial viability compared to non-irradiated controls, with a significant increase observed at 5 and 10 mGy of X-ray radiation exposure. This suggests a stimulating effect of low-dose ionizing radiation on E. coli's viability. To explore the correlation between viability and antibiotic susceptibility, we assessed the inhibition zone diameters for various antibiotics in non-irradiated and irradiated samples. The obtained results showed that the exposure of bacteria to low-dose ionizing radiation resulted in a significant reduction in the inhibition zone diameters for marbofloxacin, amoxicillin/clavulanic acid, ceftiofur, and cefoxitin. These findings suggest that low-dose X-ray radiation exposure can enhance E. coli's viability and its ability to withstand antibiotics, raising potential concerns.
Citation: Jaouhra Cherif, Anis Raddaoui, Ghofrane Ben Fraj, Asma Laabidi, Nada Souissi. Escherichia coli's response to low-dose ionizing radiation stress[J]. AIMS Biophysics, 2024, 11(2): 130-141. doi: 10.3934/biophy.2024009
Low-dose ionizing radiation can trigger a phenomenon known as hormesis in microorganisms, in which exposure to mild stressors like radiation results in beneficial adaptive responses. This study investigated the impact of low-dose X-rays on Escherichia coli's viability and their potential influence on antibiotic susceptibility. The irradiated samples displayed increased bacterial viability compared to non-irradiated controls, with a significant increase observed at 5 and 10 mGy of X-ray radiation exposure. This suggests a stimulating effect of low-dose ionizing radiation on E. coli's viability. To explore the correlation between viability and antibiotic susceptibility, we assessed the inhibition zone diameters for various antibiotics in non-irradiated and irradiated samples. The obtained results showed that the exposure of bacteria to low-dose ionizing radiation resulted in a significant reduction in the inhibition zone diameters for marbofloxacin, amoxicillin/clavulanic acid, ceftiofur, and cefoxitin. These findings suggest that low-dose X-ray radiation exposure can enhance E. coli's viability and its ability to withstand antibiotics, raising potential concerns.

| [1] |
Freneau A, Dos Santos M, Voisin P, et al. (2018) Relation between DNA double-strand breaks and energy spectra of secondary electrons produced by different X-ray energies. Inter J Radiat Biol 94: 1075-1084. https://doi.org/10.1080/09553002.2018.1518612 
|
| [2] |
Rappole CA, Mitra K, Wen H (2012) Dynamic fluorescence imaging of the free radical products of X-ray absorption in live cells. Opt Nano 1: 5. https://doi.org/10.1186/2192-2853-1-5
|
| [3] |
Chang RL, Stanley JA, Robinson MC, et al. (2020) Protein structure, amino acid composition and sequence determine proteome vulnerability to oxidation-induced damage. EMBO J 39: e104523. https://doi.org/10.15252/embj.2020104523
|
| [4] |
Courtade-Saïdi M (2007) The biological effects of very low-doses of ionizing radiation at the occupational exposure level. Morphologie 91: 166-172. https://doi.org/10.1016/j.morpho.2007.10.008
|
| [5] |
Liu F, Li N, Zhang Y (2023) The radioresistant and survival mechanisms of deinococcus radiodurans. Radiat Med Prot 4: 70-79. https://doi.org/10.1016/j.radmp.2023.03.001
|
| [6] |
Islam MT (2017) Radiation interactions with biological systems. Int J Radiat Biol 93: 487-493. https://doi.org/10.1080/09553002.2017.1286050
|
| [7] |
Calabrese EJ (2018) Hormesis: path and progression to significance. Int J Mol Sci 19: 2871. https://doi.org/10.3390/ijms19102871
|
| [8] |
Shibamoto Y, Nakamura H (2018) Overview of biological, epidemiological, and clinical evidence of radiation hormesis. Int J Mol Sci 19: 2387. https://doi.org/10.3390/ijms19082387
|
| [9] |
Lau YS, Chew MT, Alqahtani A, et al. (2021) Low dose ionising radiation-induced hormesis: therapeutic implications to human health. Appl Sci 11: 8909. https://doi.org/10.3390/app11198909
|
| [10] |
Rozhko TV, Nogovitsyna EI, Badun GA, et al. (2019) Reactive oxygen species and low-dose effects of tritium on bacterial cells. J Environ Radioact 208–209: 106035. https://doi.org/10.1016/j.jenvrad.2019.106035
|
| [11] |
Min J, Lee CW, Gu MB (2003) Gamma-radiation dose-rate effects on DNA damage and toxicity in bacterial cells. Radiat Environ Biophys 42: 189-192. https://doi.org/10.1007/s00411-003-0205-8
|
| [12] | Doss M (2013) Linear no-threshold model vs. radiation hormesis. Dose Response 24: 480-497. https://doi.org/10.2203/dose-response.13-005.Doss |
| [13] |
Broome EJ, Brown DL, Mitchel REJ (1999) Adaptation of human fibroblasts to radiation alters biases in DNA repair at the chromosomal level. Int J Radiat Biol 75: 681-690. https://doi.org/10.1080/095530099140014
|
| [14] |
Eken A, Aydin A, Erdem O, et al. (2012) Induced antioxidant activity in hospital staff occupationally exposed to ionizing radiation. Int J Radiat Biol 89: 648-653. https://doi.org/10.3109/09553002.2012.702295
|
| [15] |
Szumiel I (2012) Radiation hormesis: autophagy and other cellular mechanisms. Int J Radiat Biol 88: 619-628. https://doi.org/10.3109/09553002.2012.699698
|
| [16] |
Tubiana M, Feinendegen LE, Yang C, et al. (2009) The linear-non-threshold relationship is inconsistent with radiation biologic and experimental data. Radiology 251: 13-22. https://doi.org/10.1148/radiol.2511080671
|
| [17] |
Mullenders L, Atkinson M, Paretzke H, et al. (2009) Assessing cancer risks at low-dose radiation. Nat Rev Cancer 9: 596-604. https://doi.org/10.1038/nrc2677
|
| [18] |
Khan AUH, Blimkie M, Yang DS, et al. (2021) Effects of chronic low-dose internal radiation on immune-stimulatory responses in mice. Int J Mol Sci 7: 7303. https://doi.org/10.3390/ijms22147303
|
| [19] |
Rashid AM, Ramalingam L, Al-Ja wadi A, et al. (2018) Low dose radiation, inflammation, cancer and chemoprevention. Int J Radiat Biol 95: 506-515. https://10.1080/09553002.2018.1484194
|
| [20] | Castillo H, Smith GB (2017) Below-background ionizing radiation as an environmental cue for bacteria. Front Microbiol 8: 117. https://doi.org/10.3389/fmicb.2017.00177 |
| [21] |
Lampe N, Marin P, Coulon M, et al. (2019) Reducing the ionizing radiation background does not significantly affect the evolution of Escherichia coli populations over 500 generations. Sci Rep 17: 14891. https://doi.org/10.1038/s41598-019-51519-9
|
| [22] |
Lu YY, Fu Y, Chen LT, et al. (2023) Combined effect of simulated microgravity and low-dose ionizing radiation on structure and antibiotic resistance of a synthetic community model of bacteria isolated from spacecraft assembly room. Life Sci Space Res 38: 29-38. https://doi.org/10.1016/j.lssr.2023.04.005
|
| [23] |
Bauer AW, Kirby WMM, Sherris JC, et al. (1966) Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol 45: 493-496. https://doi.org/10.1093/ajcp/45.4_ts.493
|
| [24] | CA-SFM, Comité de l'Antibiogramme de la Société Française de Microbiologie, Recommandations vétérinaires, Paris, France, 2021. Available from: https://www.sfm-microbiologie.org/2021/12/10/casfm-veterinaire-2021/ |
| [25] |
Oskouee S, Feghhi SAH, Soleimani N (2020) Antibiotic susceptibility variations of Methicillin-resistant Staphylococcus aureus after gamma irradiation. Intern J Radiat Biol 96: 390-393. https://doi.org/10.1080/09553002.2020.1704301
|
| [26] |
Kolesnik OV, Rozhko TV, Kudryasheva NS (2023) Marine bacteria under low-intensity radioactive exposure: model experiments. Int J Mol Sci 24: 410. https://doi.org/10.3390/ijms24010410
|
| [27] |
Planel H, Soleilhavoup JP, Tixador R, et al. (1987) Influence on cell proliferation of background radiation or exposure to very low, chronic gamma radiation. Health Phys 52: 571-578. https://doi.org/10.1097/00004032-198705000-00007
|
| [28] |
Segatore B, Setacci D, Bennato F, et al. (2012) Evaluations of the effects of extremely low-frequency electromagnetic fields on growth and antibiotic susceptibility of Escherichia coli and Pseudomonas aeruginosa. Int J Microbiol 2012: 587293. https://doi.org/10.1155/2012/587293
|
| [29] |
Gottesman S (2019) Trouble is coming: signaling pathways that regulate general stress responses in bacteria. J Biol Chem 294: 11685-11700. https://doi.org/10.1074/jbc.REV119.005593
|
| [30] |
Kazmierczak MJ, Wiedmann M, Boor KJ (2005) Alternative sigma factors and their roles in bacterial virulence. Microbiol Mol Biol Rev 69: 527-543. https://doi.org/10.1128/mmbr.69.4.527-543.2005
|
| [31] |
Vaiserman A, Cuttler JM, Socol Y (2021) Low-dose ionizing radiation as a hormetin: experimental observations and therapeutic perspective for age-related disorders. Biogerontology 22: 145-164. https://doi.org/10.1007/s10522-020-09908-5
|
| [32] |
Milkovic L, Cipak Gasparovic A, Cindric M, et al. (2019) Short overview of ROS as cell function regulators and their implications in therapy concepts. Cells 8: 793. https://doi.org/10.3390/cells8080793
|
| [33] |
Huang MZ, Li JY (2020) Physiological regulation of reactive oxygen species in organisms based on their physicochemical properties. Acta Physiol 228: e13351. https://doi.org/10.1111/apha.13351
|
| [34] |
Kim BM, Hong Y, Lee S (2015) Therapeutic implications for overcoming radiation resistance in cancer therapy. Int J Mol Sci 16: 26880-26913. https://doi.org/10.3390/ijms161125991
|
| [35] |
Cherif J, Raddaoui A, Trabelsi M, et al. (2023) Diagnostic low-dose X-ray radiation induces fluoroquinolone resistance in pathogenic bacteria. Int J Radiat Biol 99: 1971-1977. https://doi.org/10.1080/09553002.2023.2232016
|
| [36] |
Li HB, Hou AM, Chen TJ, et al. (2021) Decreased antibiotic susceptibility in Pseudomonas aeruginosa surviving UV irradition. Front Microbiol 12: 604245. https://doi.org/10.3389/fmicb.2021.604245
|
| [37] |
Fang FC, Frawley ER, Tapscott T, et al. (2016) Bacterial stress responses during host infection. Cell Host Microbe 20: 133-143. https://doi.org/10.1016/j.chom.2016.07.009
|
| [38] |
Sheikh SW, Ali A, Ahsan A, et al. (2021) Insights into emergence of antibiotic resistance in acid-adapted enterohaemorrhagic Escherichia coli. Antibiotics 10: 522. https://doi.org/10.3390/antibiotics10050522
|
| [39] |
Lukačišinová M, Novak S, Paixão T (2017) Stress-induced mutagenesis: Stress diversity facilitates the persistence of mutator genes. PLoS Comput Biol 13: e1005609. https://doi.org/10.1371/journal.pcbi.1005609
|
| [40] |
Tahmasebi H, Dehbashi S, Arabestani MR (2021) Antibiotic resistance alters through iron-regulating sigma factors during the interaction of Staphylococcus aureus and Pseudomonas aeruginosa. Sci Rep 11: 18509. https://doi.org/10.1038/s41598-021-98017-5
|
| [41] |
Maurya N, Jangra M, Tambat R, et al. (2019) Alliance of efflux pumps with beta-Lactamases in multidrug-resistant Klebsiella pneumoniae isolates. Microb Drug Resist 25: 1155-1163. https://doi.org/10.1089/mdr.2018.0414
|
| [42] |
Zhao L, Wang S, Li X, et al. (2020) Development of in vitro resistance to fluoroquinolones in Pseudomonas aeruginosa. Antimicrob. Resist Infect Control 9: 124. https://doi.org/10.1186/s13756-020-00793-8
|
| [43] |
Ghai I (2023) Barrier to entry: examining the bacterial outer membrane and antibiotic resistance. Appl Sci 13: 4238. https://doi.org/10.3390/app13074238
|
| [44] |
Nagashima H, Shiraishi K, Ohkawa S, et al. (2018) Induction of somatic mutations by low-dose X-rays: the challenge in recognizing radiation-induced events. J Radiat Res 59: 11-17. https://doi.org/10.1093/jrr/rrx053
|
| [45] |
Davin-Regli A, Pages JM, Ferrand A (2021) Clinical status of efflux resistance mechanisms in gram-negative bacteria. Antibiotics (Basel) 10: 1117. https://doi.org/10.3390/antibiotics10091117
|
| [46] |
Li S, Li Z, Wei W, et al. (2015) Association of mutation patterns in GyrA and ParC genes with quinolone resistance levels in lactic acid bacteria. J Antibiot 68: 81-87. https://doi.org/10.1038/ja.2014.113
|
| [47] |
Redgrave LS, Sutton SB, Webber MA, et al. (2014) Fluoroquinolone resistance: mechanisms, impact on bacteria, and role in evolutionary success. Trends Microbiol 22: 438-445. https://doi.org/10.1016/j.tim.2014.04.007
|
Figures(2) / Tables(2)

Jaouhra Cherif, Anis Raddaoui, Ghofrane Ben Fraj, Asma Laabidi, Nada Souissi. Escherichia coli's response to low-dose ionizing radiation stress[J]. AIMS Biophysics, 2024, 11(2): 130-141. doi: 10.3934/biophy.2024009







 DownLoad:
DownLoad: