Citation: Charles O. Nwamba, Ferdinand C. Chilaka, Ali Akbar Moosavi-Movahedi. Cation modulation of hemoglobin interaction with sodium n-dodecyl sulphate (SDS) iv: magnesium modulation at pH 7.20[J]. AIMS Biophysics, 2016, 3(1): 146-170. doi: 10.3934/biophy.2016.1.146

| [1] | Kumar S, Ma B, Tsai CJ, et al. (2000) Folding and binding cascades: dynamic landscapes and population shifts. Protein Sci 9: 10–19. |
| [2] |
Zhuravlev PI, Papoian GA (2010). Functional versus folding landscapes: the same yet different. Curr Opin Struct Biol 20: 16–22. doi: 10.1016/j.sbi.2009.12.010

|
| [3] |
Del Sol A, Tsai C-J, Ma B, et al. (2009). The Origin of Allosteric Functional Modulation: Multiple Pre-existing Pathways. Structure 17: 1042–1050. doi: 10.1016/j.str.2009.06.008
|
| [4] |
Kar G, Keskin O, Gursoy A, et al. (2010). Allostery and population shift in drug discovery. Curr Opin Pharmacol 10: 715–722. doi: 10.1016/j.coph.2010.09.002
|
| [5] | Poillon WN, Kim BC (1990) 2,3-Diphosphoglycerate and intracellular pH as interdependent determinants of the physiologic solubility of deoxyhemoglobin S. Blood 76: 1028–1036. |
| [6] | Nadolny C, Kempf I, Zundel G (1993) Specific interactions of the allosteric effector 2,3-bisphosphoglycerate with human hemoglobin-a difference FTIR study. Biolog Chem Hoppe Seyler 374(6): 403–407. |
| [7] |
Low C, Homeyer N, Weininger U, et al. (2009) Conformational switch upon phosphorylation: human CDK inhibitor p19INK4d between the native and partially folded state. ACS Chem Biol 4: 53–63. doi: 10.1021/cb800219m
|
| [8] | Allison AC (2002). The discovery of resistance to malaria of sickle-cell heterozygotes. In mini-series: significant contributions to biological chemistry over the past 125 years. Biochem Mol Biol Edu 30: 279–287. |
| [9] | Bonaventura C, Henkens R, Alayash AI, et al. (2013) Molecular Controls of the Oxygenation and Redox Reactions of Hemoglobin. Antioxid Redox Signal 18 (17): 2298–2313. |
| [10] | Chilaka FC, Nwamba CO, Moosavi-Movahedi AA (2011) Cation Modulation of Hemoglobin Interaction with Sodium n-Dodecyl Sulfate (SDS). I: Calcium Modulation at pH 7.20. Cell Biochem Biophy 60: 187–197. |
| [11] | Nwamba CO, Chilaka FC, Moosavi-Movahedi AA (2011) Cation Modulation of Hemoglobin Interaction with Sodium n-Dodecyl Sulfate (SDS). II: Calcium Modulation at pH 5.0, Cell Biochem Biophy 61: 573–584. |
| [12] | Nwamba CO, Chilaka FC, Moosavi-Movahedi AA (2013) Cation Modulation of Hemoglobin Interaction with Sodium n-Dodecyl Sulfate (SDS). III: Calcium Modulation at pH 5.0, Cell Biochem Biophy 67: 547–555. |
| [13] |
Kelemen C, Chien S, Artmann GM (2001) Temperature transition of human hemoglobin at body temperature: effects of calcium. Biophysical J 80: 2622–2630. doi: 10.1016/S0006-3495(01)76232-7
|
| [14] | Swaminathan R. (2003) Magnesium Metabolism and its Disorders. Clin Biochem Rev 24: 47–66. |
| [15] | Luft, FC (2012) Whither Magnesium. Clin Kidney J 5(Suppl 1): i1–i2. |
| [16] |
Altura BM, Altura BT, Gebrewold A, et al. (1984) Magnesium deficiency and hypertension: correlation between magnesium-deficient diets and microcirculatory changes in situ. Science 223: 1315–1317. doi: 10.1126/science.6701524
|
| [17] |
Laurant P, Hayoz D, Brunner HR, et al. (1999) Effect of magnesium deficiency on blood pressure and mechanical properties of rat carotid artery. Hypertension 33: 1105–1110. doi: 10.1161/01.HYP.33.5.1105
|
| [18] |
Laires, MJ, Monteiro CP, Bicho M (2004) Role of cellular magnesium in health and human disease. Frontiers Biosc. 9: 262–276. doi: 10.2741/1223
|
| [19] |
Olson JA, Kilejian A (1982) Involvement of spectrin and ATP erythrocyte ghosts by the human Plasmodium falciparum. J Cell Biol 95: 757–762. doi: 10.1083/jcb.95.3.757
|
| [20] | Atamna H, Ginsburg H (1997) The malaria parasite supplies glutathione to its host cell. Investigation of glutathione transport and metabolism in human eryhtrocytes infected with Plasmodium falciparum. Eur J Biochem 250: 670–679. |
| [21] | Mauritz JMA, Esposito A, Ginsburg H, et al. (2009) The Homeostasis of Plasmodium falciparum-Infected Red Blood Cells. PLoS Comput Biol 5(4): e1000339. |
| [22] |
Hanada K, Mitamura T, Fukasawa M, et al. (2000). Neutral sphingomyelinase activity dependent on Mg2+ and anionic phospholipids in the intraerythrocytic malaria parasite Plasmodium falciparum. Biochemical J 346: 671–677. doi: 10.1042/bj3460671
|
| [23] |
Labaied M, Dagan A, Dellinger M, et al. (2004). Anti-Plasmodium activity of ceramide analogs. Malaria J 3: 49–58. doi: 10.1186/1475-2875-3-49
|
| [24] | Wester PO (1987) Magnesium. Am J Clin Nutr 45: 1305–1312. |
| [25] | Kujaník Š (2003) Magnesium and the treatment of some cardiovascular diseases. Acta Medica Martiniana 3: 3–8. |
| [26] |
Lew VL, Bookchin RM (2005) Ion transport pathology in the mechanism of sickle cell dehydration. Physiol Rev 85: 179–200. doi: 10.1152/physrev.00052.2003
|
| [27] |
Aprelev A, Rotter MA, Etzion Z, et al. (2005). The effects of erythrocyte membranes on the nucleation of sickle hemoglobin. Biophysical J 88: 2815–2822. doi: 10.1529/biophysj.104.051086
|
| [28] |
Gibson JS, Khan A, Speake PF et al. (2001). O2 dependence of K+ transport in sickle cells: the effect of different cell populations and the substituted benzaldehyde 12C79. FASEB J 15: 823–832. doi: 10.1096/fj.00-0177com
|
| [29] |
Lew VL, Etzion Z, Bookchin RM (2002) Dehydration response of sickle cells to sickling-induced Ca2+ permeabilization. Blood 99: 2578–2585. doi: 10.1182/blood.V99.7.2578
|
| [30] |
Maher AD, Kuchel PW (2003) The Gárdos channel: a review of the Ca2+-activated K+ channel in human erythrocytes. The Intl J Biochem Cell Biol 35: 1182–1197. doi: 10.1016/S1357-2725(02)00310-2
|
| [31] | Ward GE, Miller LH, Dvorak JA (1993) The origin of parasitophorous vacuole membrane lipids in malaria-infected erythrocytes. J Cell Sci 106: 237–248. |
| [32] | Dluzewski AR, Zicha D, Dunn GA, et al. (1995) Origins of the parasitophorous vacuole membrane of the malaria parasite: surface area of the parasitized red cell. Euro J Cell Biol 68: 446–449. |
| [33] |
Murphy SC, Samuel BU, Harrison T, et al. (2004). Erythrocyte detergent-resistant membrane proteins: their characterization and selective uptake during malarial infection. Blood 103: 1920–1928. doi: 10.1182/blood-2003-09-3165
|
| [34] | Kirk K (2001). Membrane transport in the malaria-infected erythrocyte. Physiol Rev, 81: 495–537. |
| [35] | Banerjee R, Goldberg DE (2000) The Plasmodium food vacuole. Antimalarial Chemotherapy: Mechanisms of Action, Resistance, and New Directions in Drug Discovery. P.J. Rosenthal, editor. Humana Press, Totowa, NJ. 43–63. |
| [36] | Klemba M, Beatty W, Gluzman I, et al. (2004). Trafficking of plasmepsin II to the food vacuole of the malaria parasite Plasmodium falciparum. The J Cell Biol 164 (1): 47–56. |
| [37] |
Tekwani BL, Walker LA (2005). Targeting the hemozoin synthesis pathway for new antimalarial drug discovery: technologies for in vitro β-hematin formation assay. Combinat Chem High Throughput Screening, 8: 63–79. doi: 10.2174/1386207053328101
|
| [38] | Millart H, Durlach V, Durlach J (1995) Red blood cell magnesium concentrations: analytical problems and significance. Magnesium Res 8: 65–76. |
| [39] |
Shin I, Kreimer D, Silman I, et al. (1997). Membrane-promoted unfolding of acetylcholinesterase: A possible mechanism for insertion into the lipid bilayer. PNAS 94: 2848–2852. doi: 10.1073/pnas.94.7.2848
|
| [40] |
Tanaka A, Hoshino E (2003) Similarities between the thermal inactivation kinetics of Bacillus amyloliquefaciens α-amylase in an aqueous solution of sodium dodecyl sulphate and the kinetics in the solution of anionic phospholipid vesicles. Biotechnol Appl Biochem 38: 175–181. doi: 10.1042/BA20030050
|
| [41] |
Moosavi-Movahedi AA, Nazari K, Saboury AOA (1997) Thermodynamics of denaturation of horseradish peroxidase with sodium n-dodecyl sulphate and n-dodecyl trimethylammonium bromide. Colloids and Surfaces B: Biointerfaces, 9: 123–130. doi: 10.1016/S0927-7765(97)00016-7
|
| [42] | Ajloo D, Moosavi-Movahedi AA, Sadeghi M, et al. (2002) Comparative, structural and functional studies of avian and mammalian haemoglobins. Acta Biochim Polonica 49: 459–470. |
| [43] |
Bordbar AK, Moosavi-Movahedi AA, Amini MK (2003) A microcalorimetry and binding study on interaction of dodecyl trimethylammonium bromide with wigeon haemoglobin. Thermochimica Acta 400: 95–100. doi: 10.1016/S0040-6031(02)00483-5
|
| [44] | Otzen D (2011) Protein–surfactant interactions: a tale of many states. Biochem Biophy Acta 1814: 562–591. |
| [45] |
William RC Jr, Tsay KY (1973) A convenient chromatographic method for the preparation of human hemoglobin. Anal Biochem 54: 137–145. doi: 10.1016/0003-2697(73)90256-X
|
| [46] |
Riggs A (1981) Preparation of blood hemoglobin of vertebrates. Methods Enzymol 76: 5–29. doi: 10.1016/0076-6879(81)76111-1
|
| [47] | Antonini E, Brunori M (1971). The derivatives of ferrous hemoglobin and myoglobin. In: Neuberger, A., Tatum, E. L., Eds. Hemoglobin and myoglobin in their reactions with ligands, Amsterdam: North-Holland Publishing Co. 21: 13–39. |
| [48] | Coletta M, Ascenzi P, Santucci R, et al. (1993). Interaction of inositol hexakisphosphate with liganded ferrous human hemoglobin: direct evidence for two functionally operative binding sites. Biochem Biophys Acta 1162: 309–314. |
| [49] | Schmid F-X (2001) Biological Macromolecules: UV-visible Spectrophotometry. Encyc Life Sci (eLS) 3: 240–243. |
| [50] |
Ichikawa T, Terada H (1977) Second derivative spectrophotometry as an effective tool for examining phenylalanine residues in proteins. Biochim Biophys Acta 494: 267–270. doi: 10.1016/0005-2795(77)90154-4
|
| [51] |
Ichikawa T, Terada H (1979) Estimation of state and amount of phenylalanine residues in proteins by second derivative spectrophotometry. Biochim Biophy Acta 580: 120–128. doi: 10.1016/0005-2795(79)90203-4
|
| [52] | Mach H, Middaugh CR (1994) Simultaneous monitoring of the environment of tryptophan, tyrosine, and phenylalanine residues in proteins by near-ultraviolet second-derivative spectroscopy. Anal Biochem 222(2): 323–331. |
| [53] |
Bellelli A, Brunori M (1994) Optical measurements of quaternary structural changes in hemoglobins. Methods Enzymol 232: 56–71. doi: 10.1016/0076-6879(94)32043-8
|
| [54] |
Vallone B, Bellelli A, Miele AE, et al. (1996) Probing the α12 Interface of Human Hemoglobin by Mutagenesis: Role of the FG-C Contact regions. J Biol Chem 271: 12472–12480. doi: 10.1074/jbc.271.21.12472
|
| [55] |
Fang T-Y, Simplaceanu V, Tsai C-H, et al. (2000) An additional H-bond in the α12 Interface as the structural basis for the low oxygen affinity and high cooperativity of a novel recombinant hemoglobin (L105W). Biochemistry 39: 13708–13718. doi: 10.1021/bi001115i
|
| [56] |
Tsai C-H, Ho C (2002) Recombinant hemoglobins with low oxygen affinity and high cooperativity. Biophys Chem 98: 15–25. doi: 10.1016/S0301-4622(02)00081-9
|
| [57] |
Mach H, Middaugh CR, Lewis RV (1992) Statistical determination of the average values of the extinction coefficients of tryptophan and tyrosine in native proteins. Anal Biochem 200: 74–80. doi: 10.1016/0003-2697(92)90279-G
|
| [58] |
Wetlaufer DB (1963) Ultraviolet spectra of proteins and amino acids. Adv Protein Chem 17: 303–390. doi: 10.1016/S0065-3233(08)60056-X
|
| [59] | Nwamba CO, Chilaka FC (2010) A proposed significance of the δ region and its implications in the Mechanism of Cooperativity in Hemoglobin. Medical Hypoth Res 6: 25–35. |
| [60] | Nagababu E, Rifkind J (2000). Reaction of Hydrogen Peroxide with Ferrylhemoglobin: Superoxide Production and Heme Degradation, Biochemistry, 39: 12503–12511. |
| [61] |
Salehi N, Moosavi-Movahedi AA, Fotouhi L, et al. (2014). Heme degradation upon production of endogenous hydrogen peroxide via interaction of hemoglobin with sodium dodecyl sulfate. J Photochem Photobiol B: Biology 133: 11–17. doi: 10.1016/j.jphotobiol.2014.02.014
|
| [62] |
Maitra D, Byun J, Andreana PR, et al. (2011). Mechanism of hypochlorous acid-mediated heme destruction and free iron release. Free Rad Biol Med 51: 364–373. doi: 10.1016/j.freeradbiomed.2011.03.040
|
| [63] |
Polet H, Steinhardt J (1969) Sequential stages in the acid denaturation of horse and human ferrihemoglobins. Biochemistry 8: 857–864. doi: 10.1021/bi00831a014
|
| [64] | Jarolim P, Lahav M, Liu S-C, et al. (1990) Effect of hemoglobin oxidation products on the stability of red cell membrane skeletons and the associations of skeletal proteins: correlation with a release of hemin. Blood 76: 2125–2131. |
| [65] |
Asakura T, Ohnishi T, Friedman S, et al. (1974) Abnormal precipitation of oxyhemoglobin S by mechanical shaking. PNAS 71: 1594–1598. doi: 10.1073/pnas.71.5.1594
|
| [66] |
Asakura T, Minakata K, Adachi K, et al. (1977) Denatured hemoglobin in sickle erythrocytes. J Clin Invest 59: 633–640. doi: 10.1172/JCI108681
|
| [67] | Cooper CE, Jurd M, Nicholls P, et al. (2005) On the formation, nature, stability and biological relevance of the primary reaction intermediates of myoglobins with hydrogen peroxide. Dalton Trans 2005 (21): 3483–3488. |
| [68] |
Rifkind JM, Abugo O, Levy A, et al. (1994) Detection, formation, and relevance of hemichromes and hemochromes. Methods Enzymol 231: 449–480. doi: 10.1016/0076-6879(94)31030-0
|
| [69] | Giulivi C, Davies KJA (1990) A novel antioxidant role of hemoglobin: the comproportionation of ferrylhemoglobin with oxyhemoglobin. J Biol Chem 265: 19453–19460. |
| [70] |
Giulivi C, Davies KJA (1994) Hydrogen-peroxide mediated ferrylhemoglobin generation in vitro and in red blood cells. Methods Enzymol 231: 490–496. doi: 10.1016/0076-6879(94)31032-7
|
| [71] |
Giulivi C, Cadenas E (1994) Ferrylmyoglobin: Formation and Chemical Reactivity toward Electron-Donating Compounds. Methods Enzym 233: 189–202. doi: 10.1016/S0076-6879(94)33022-0
|
| [72] |
Svistunenko DA (2005) Reaction of haem containing proteins and enzymes with hydroperoxides: The radical view. Biochim Biophys Acta 1707: 127–155. doi: 10.1016/j.bbabio.2005.01.004
|
| [73] | Yonetani T, Schleyer H (1967) Studies on Cytochrome C Peroxidase IX. The reaction of ferrimyoglobin with hydroperoxides and a comparison of peroxide-induced compounds of ferrimyoglobin and cytochrome C peroxidase. J Biol Chem 242(8): 1974–1979. |
| [74] |
Brown SB, Shillcock M, Jones P (1976) Equilibrium and kinetic studies of the aggregation of porphyrins in aqueous solution. Biochem J 153: 279–285. doi: 10.1042/bj1530279
|
| [75] | La Brake CC, Fung LW-M (1998) Sickle hemoglobin is more fusogenic than normal hemoglobin at physiological pH and ionic strength conditions. Biochem Biophys Acta 1406: 152–161. |
| [76] |
De Franceschi L, Bachir D, Galacteros F, et al. (1997) Oral Magnesium supplements reduce erythrocyte dehydration in patients with sickle cell disease. J Clin Invest 100: 1847–1852. doi: 10.1172/JCI119713
|
| [77] | Kirk K (2001) Membrane transport in the malaria-infected erythrocyte. Physiol Rev, 81: 495–537. |
| [78] |
Goldberg DE, Slater AFG, Cerami A, et al. (1990) Haemoglobin degradation in the malaria parasite, Plasmodium falciparum: An ordered process in a unique organelle. PNAS 87: 2931–2935. doi: 10.1073/pnas.87.8.2931
|
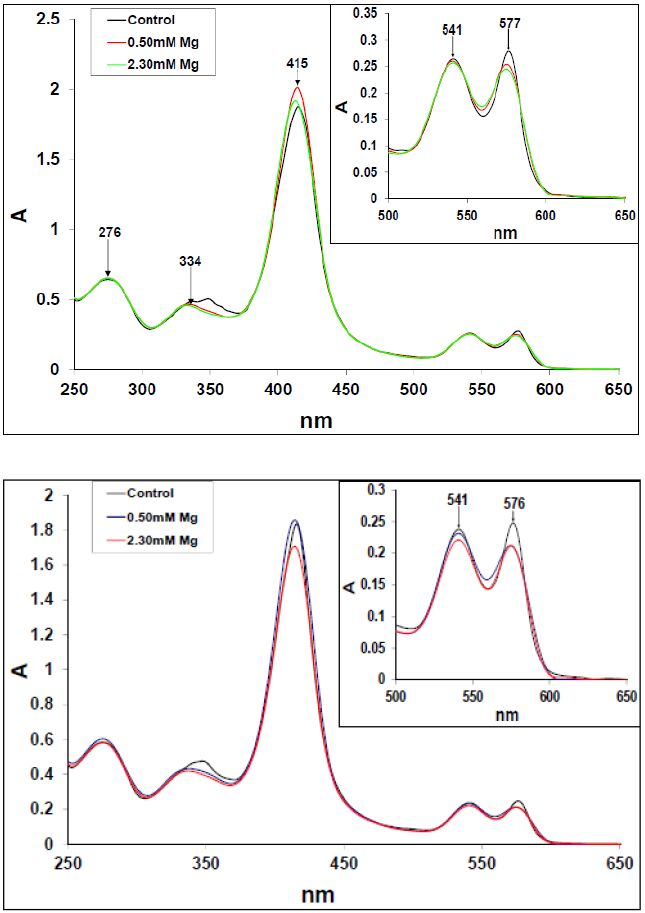
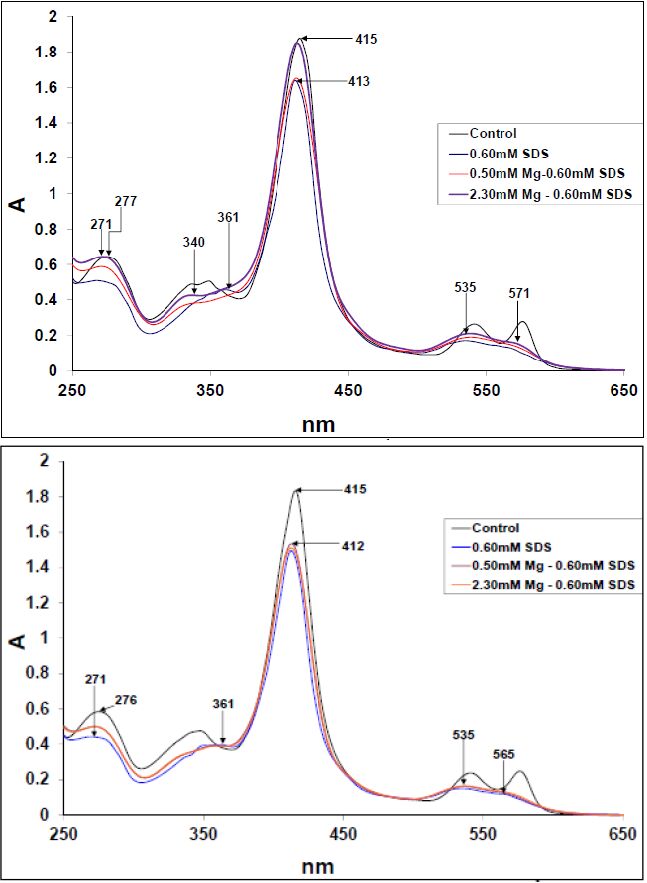
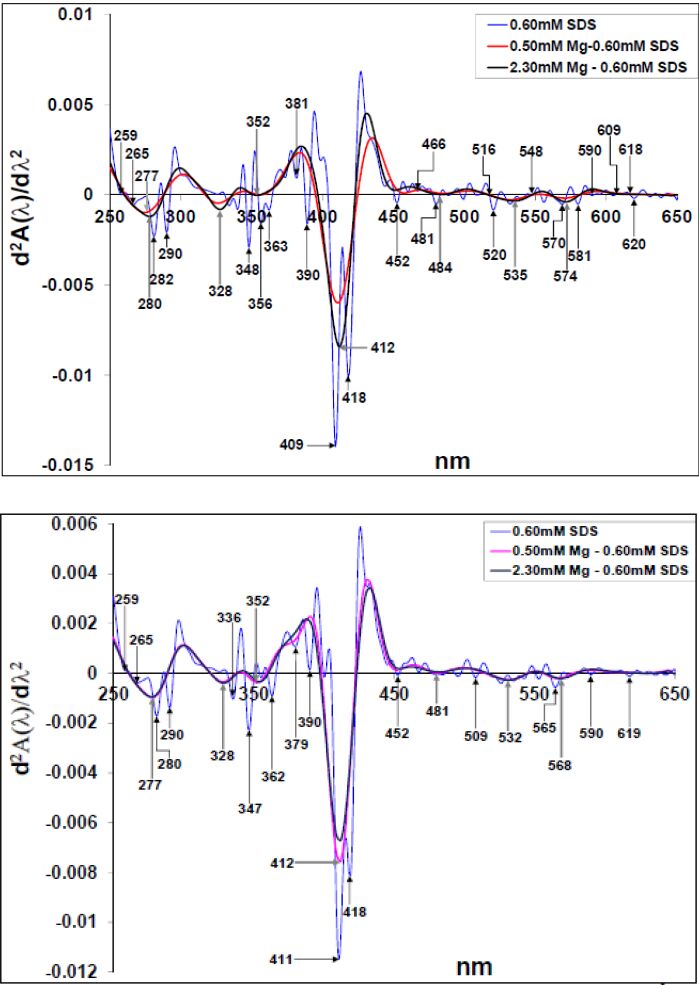
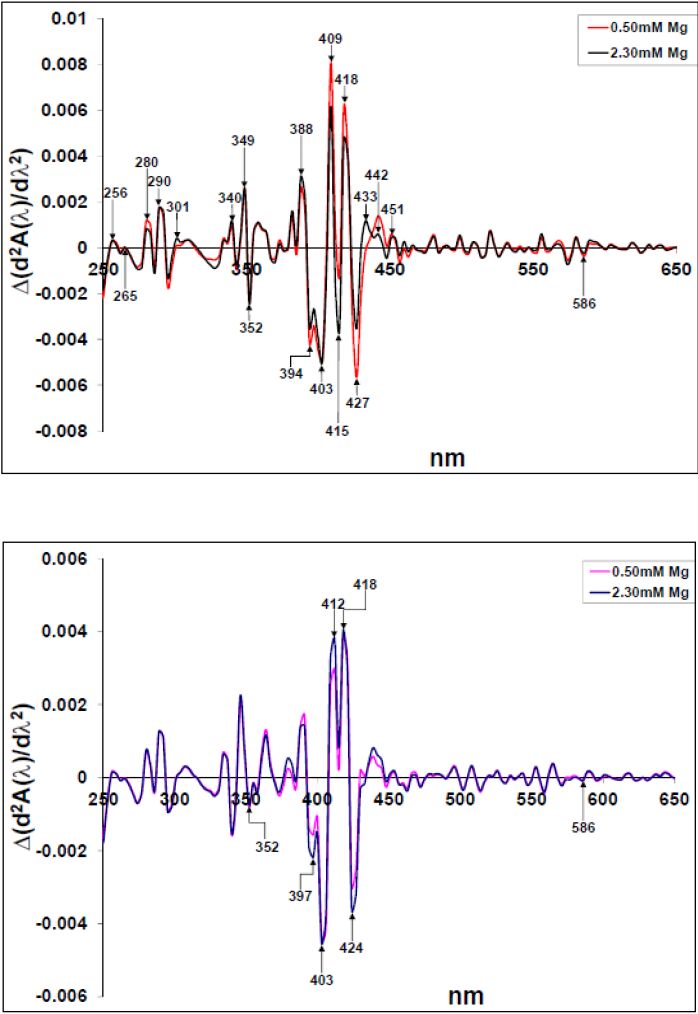
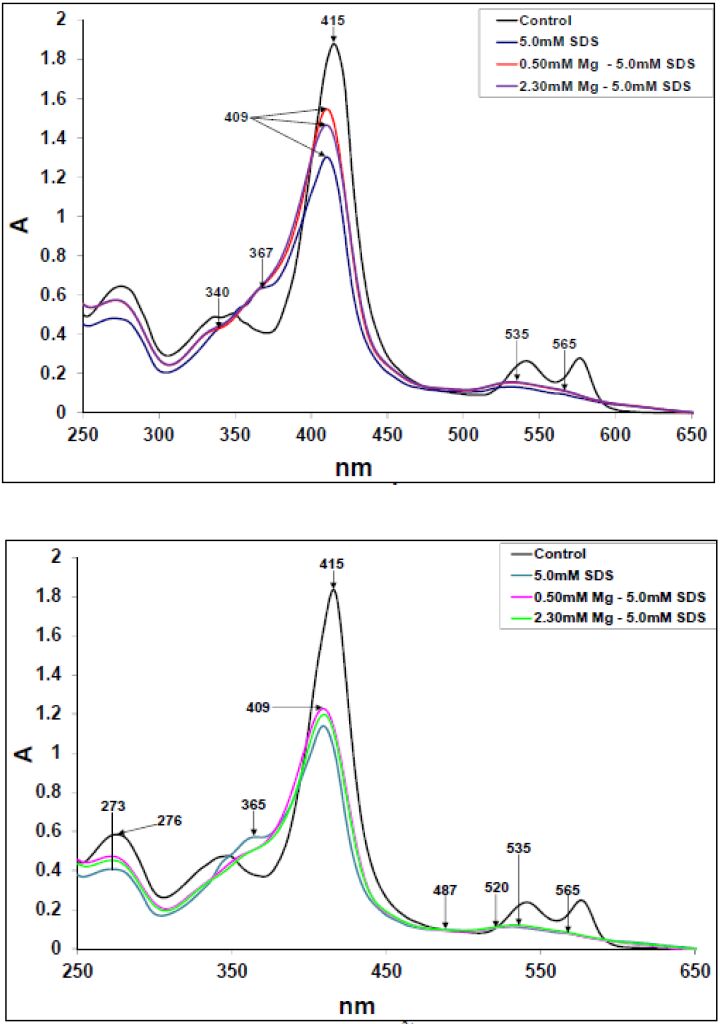
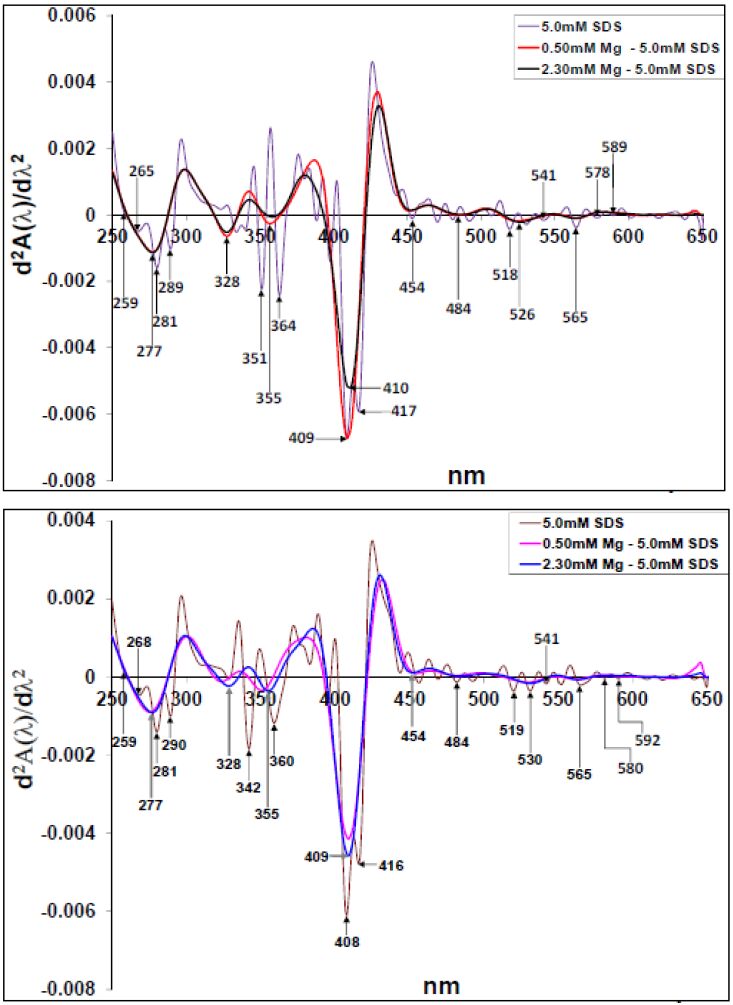
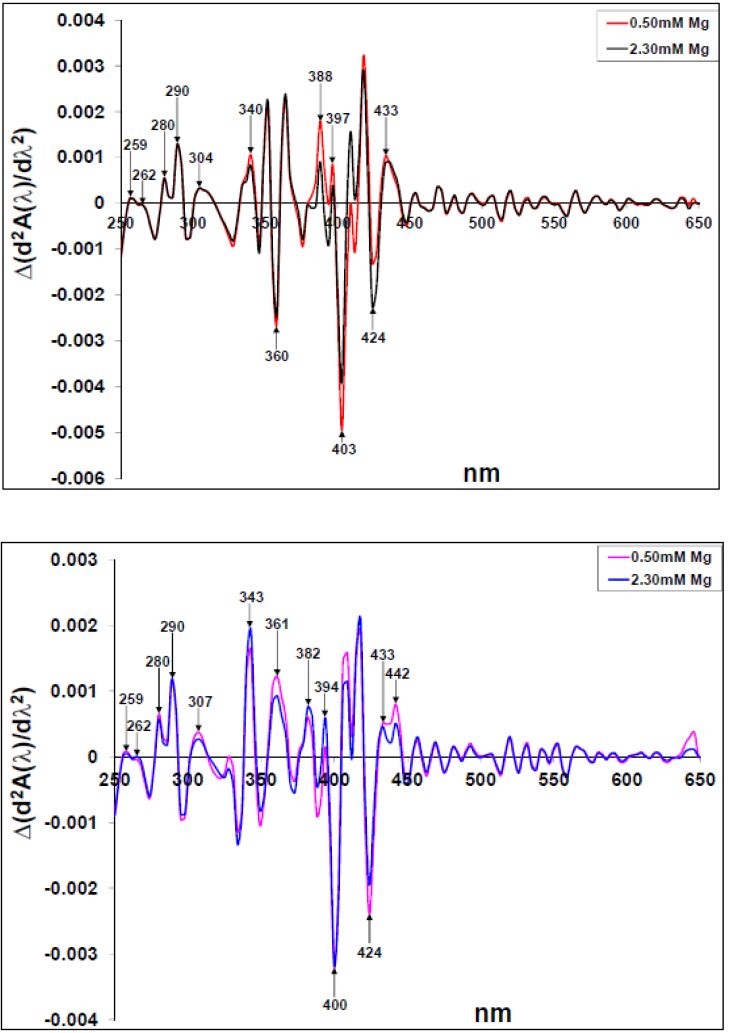
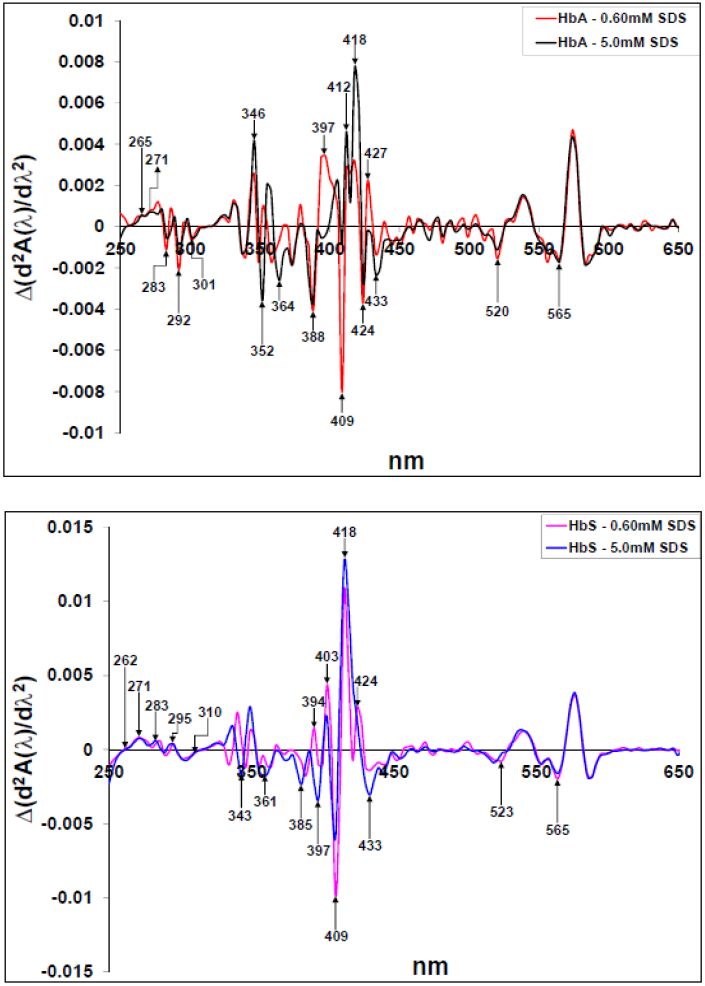
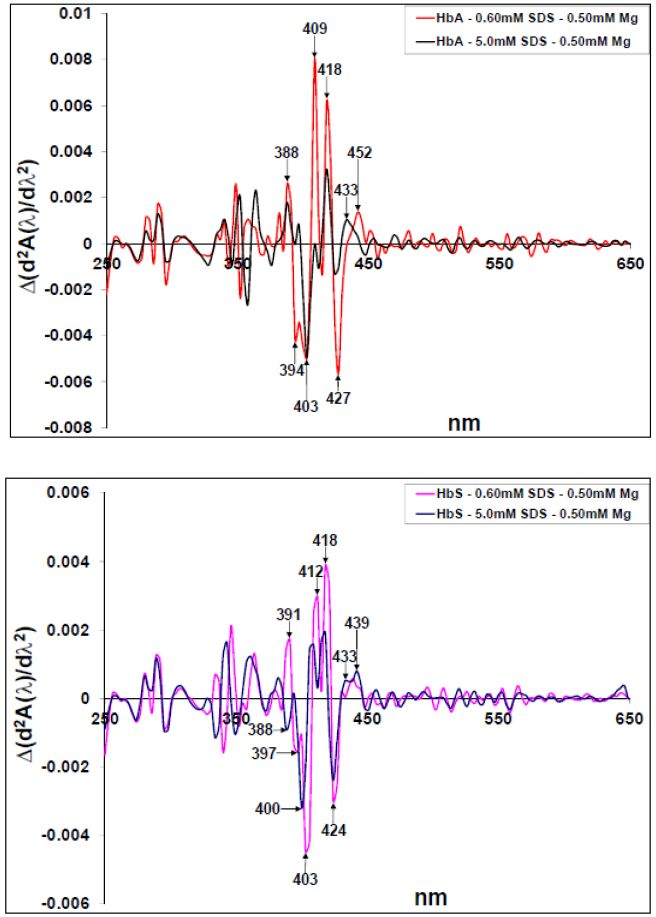
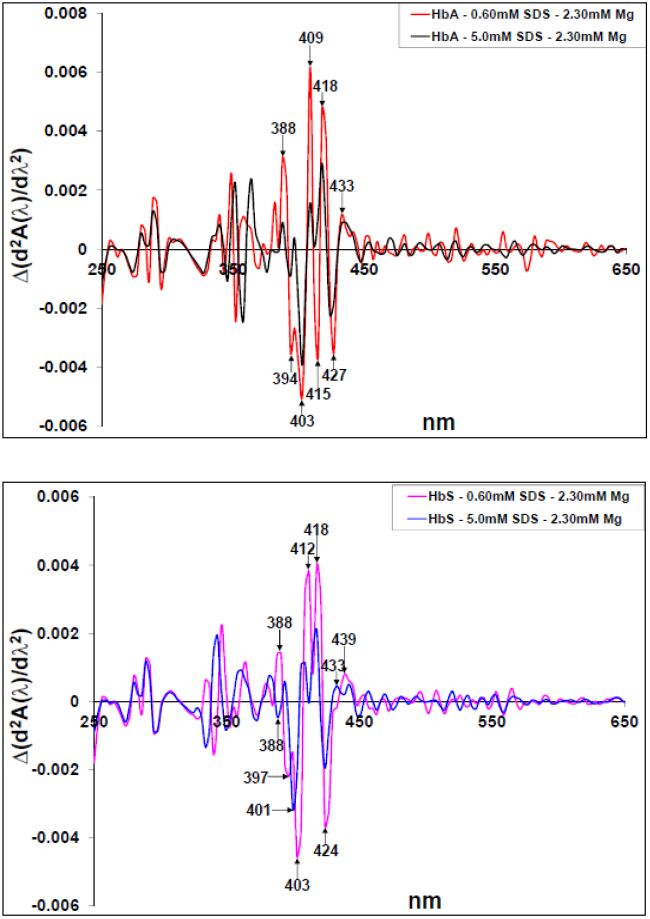
Figures(10)

Charles O. Nwamba, Ferdinand C. Chilaka, Ali Akbar Moosavi-Movahedi. Cation modulation of hemoglobin interaction with sodium n-dodecyl sulphate (SDS) iv: magnesium modulation at pH 7.20[J]. AIMS Biophysics, 2016, 3(1): 146-170. doi: 10.3934/biophy.2016.1.146







 DownLoad:
DownLoad: