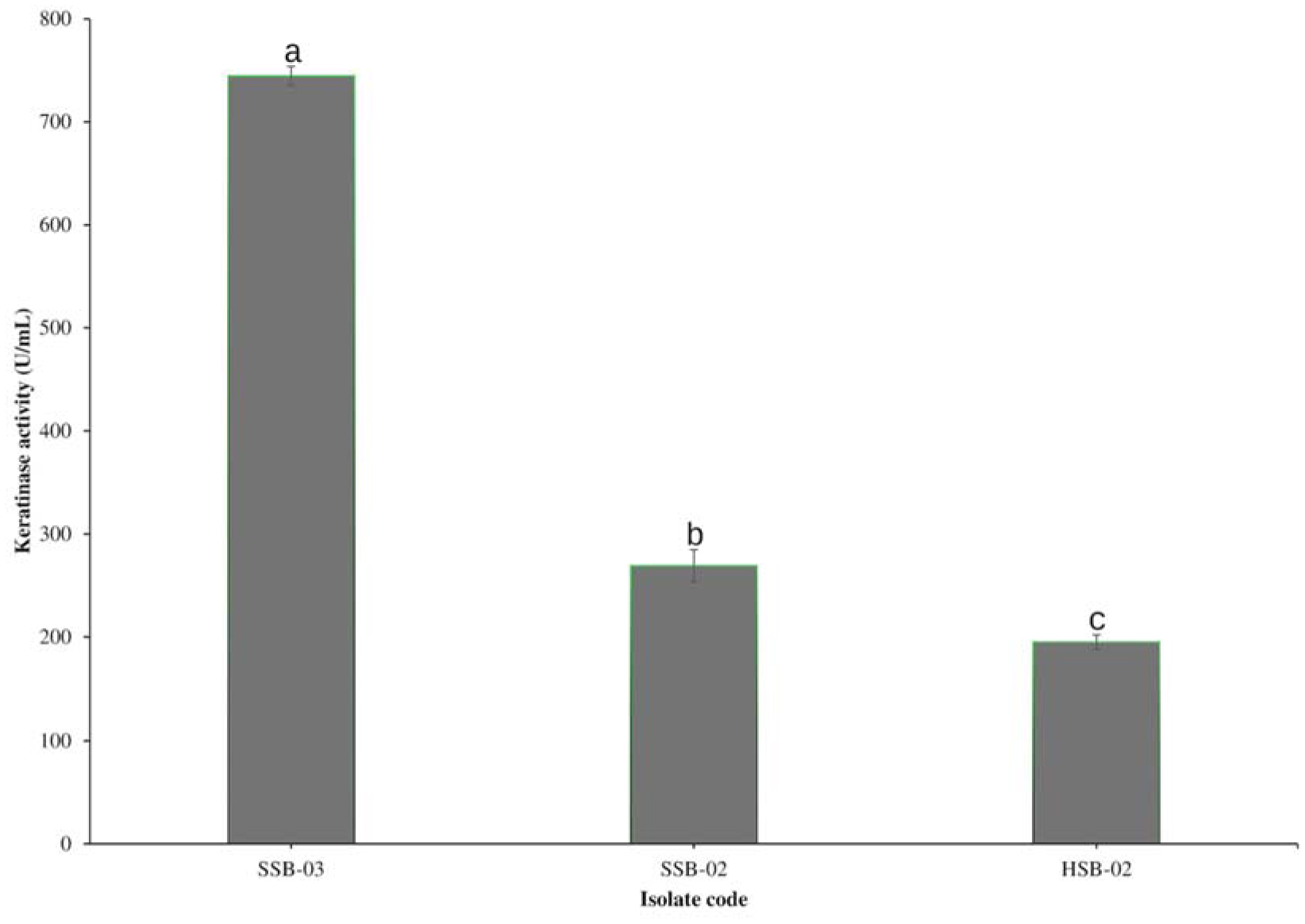
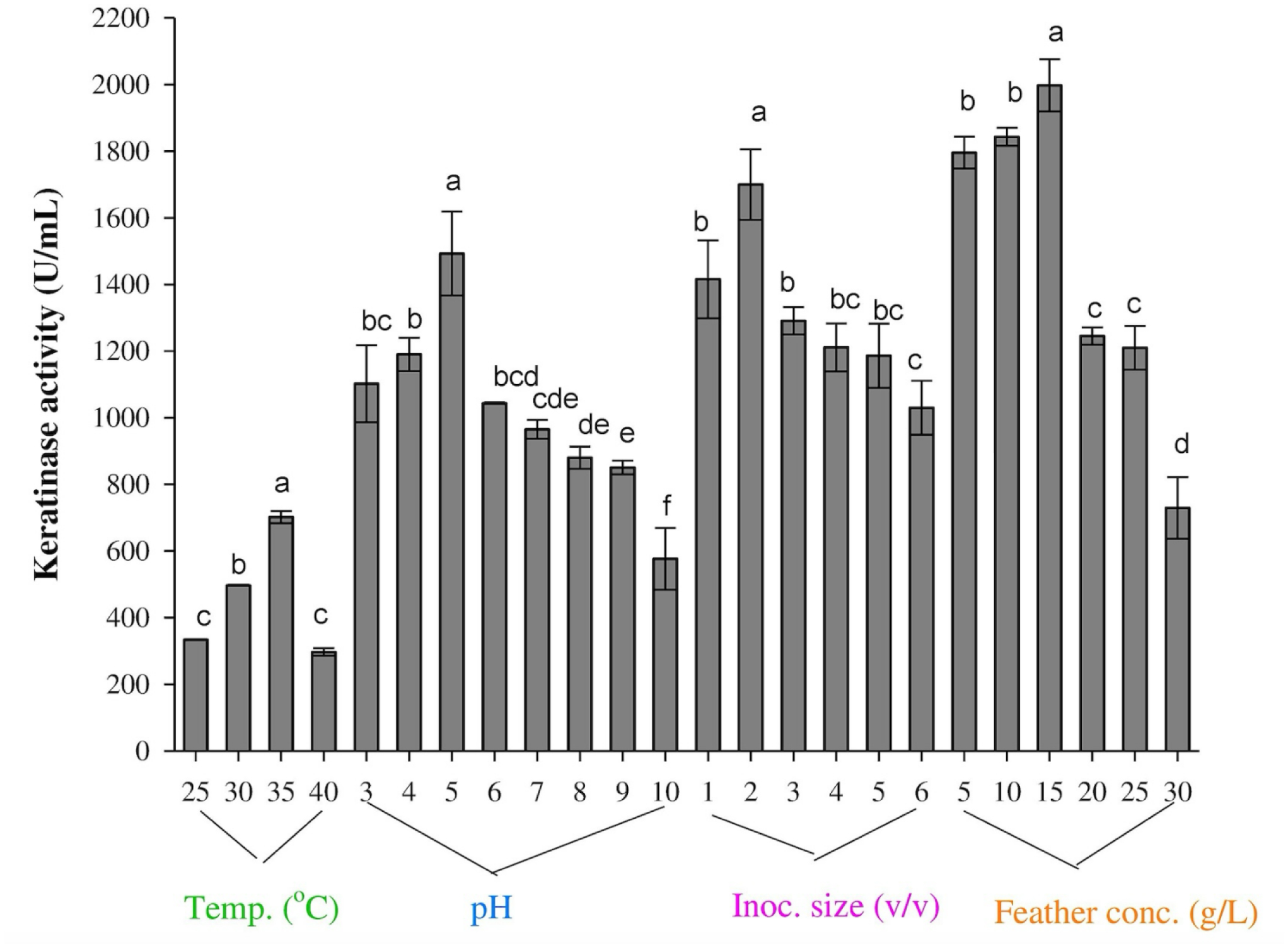
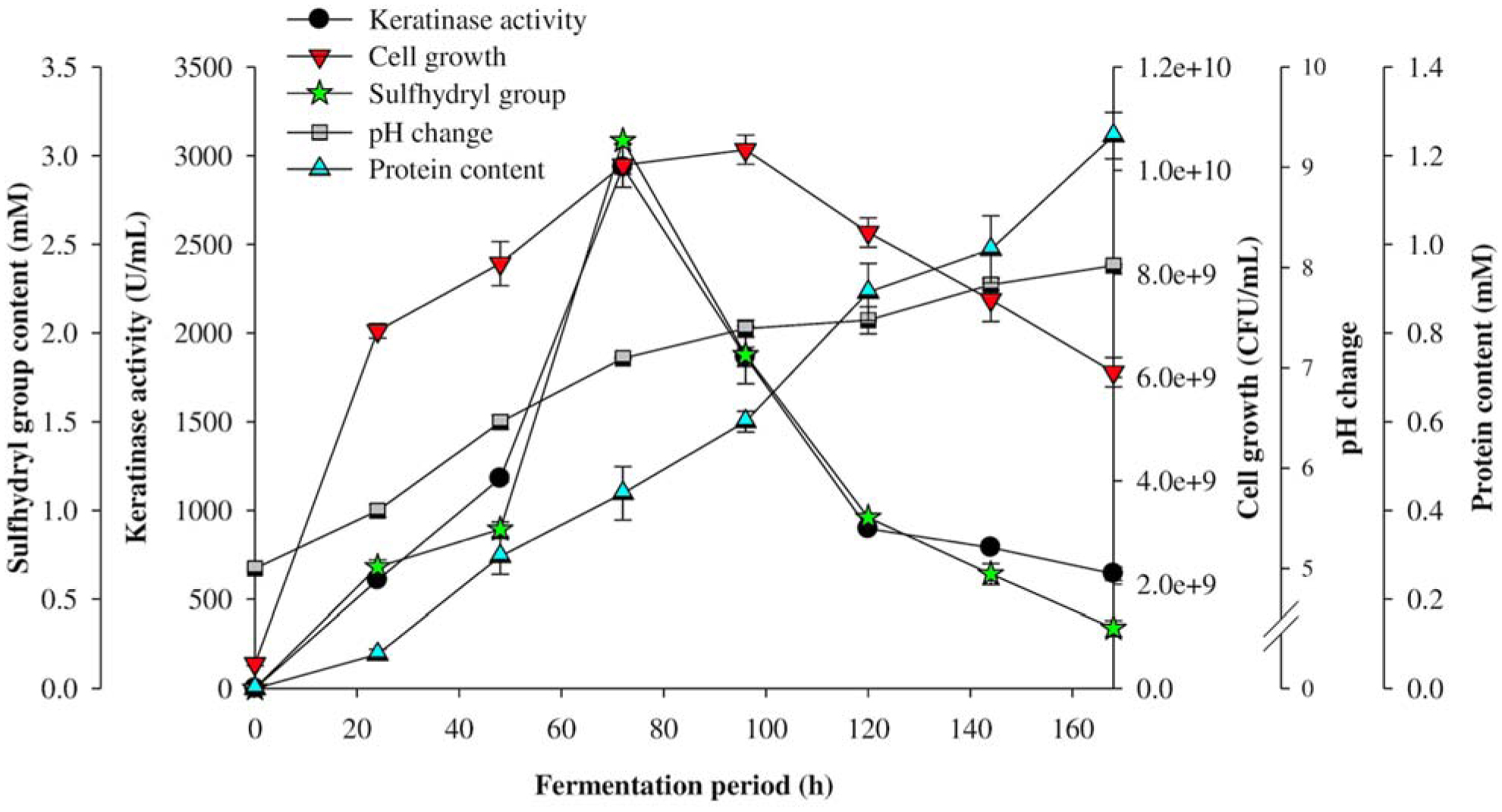
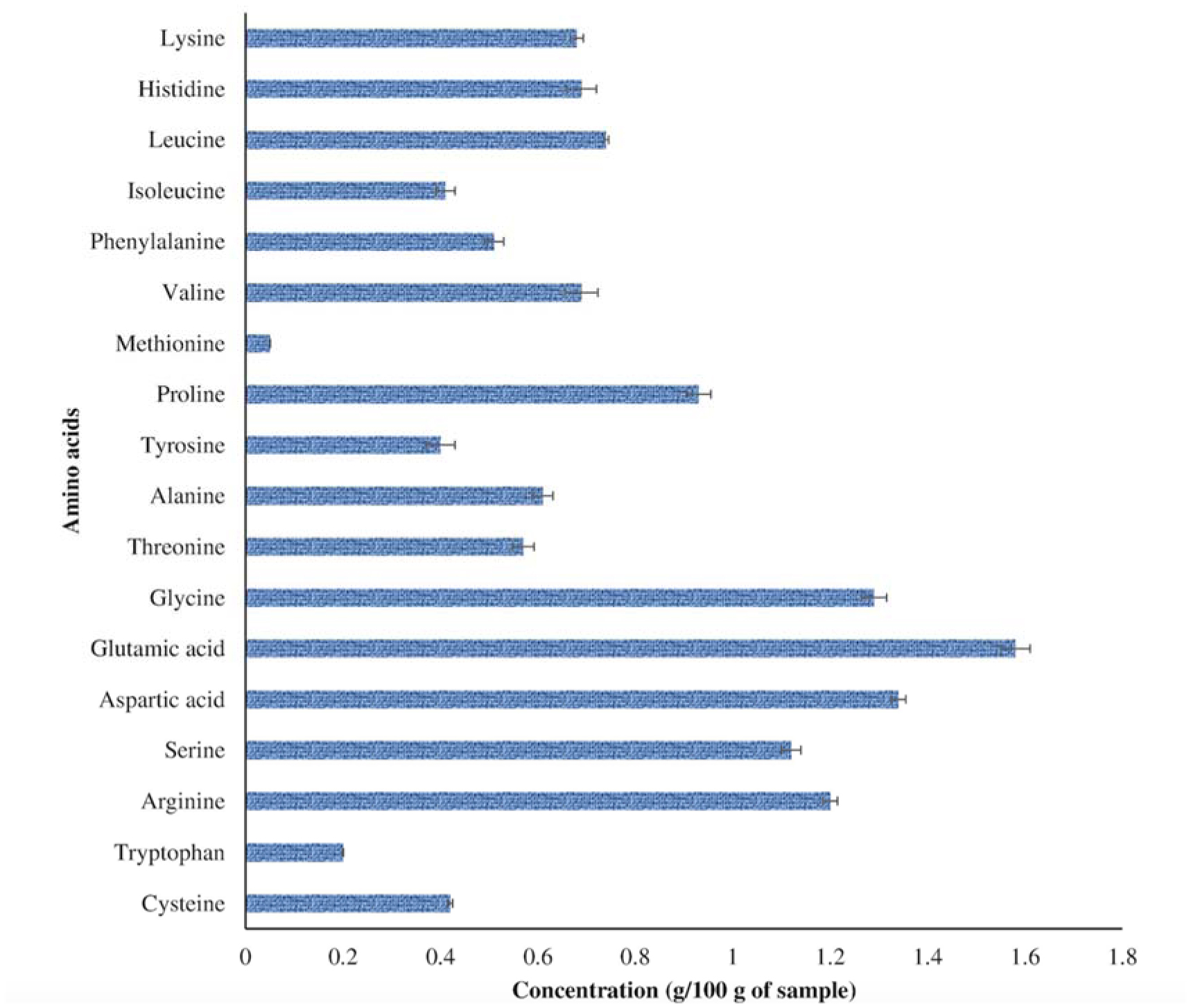
The transition from a traditional linear economy to a circular model aims to create a more sustainable future by reducing the adverse effects of agro-waste on the environment. The present study evaluated the metabolic diversity of bacterial isolates from municipal dumpsites for keratinase production and poultry feather valorization. The bacterium with the highest keratinolytic activity was identified through 16S ribosomal ribonucleic acid (rRNA) gene sequencing. The exo-keratinase production by the bacterium was optimized, and the feather hydrolysate obtained from the fermentation process was analyzed for amino acids. Among the twelve bacteria isolated from the dumpsite's sample, three showed significant feather degradation and keratinase production of 89% (744.5 ± 9.19 U/mL), 58% (269 ± 15.55 U/mL), and 46% (195 ± 7.07 U/mL) for SSB-03, SSB-02, and HSB-02, respectively. Analysis of the 16S rRNA gene sequence revealed that SSB-03 has high sequence homology with Exiguobacterium acetylicum, and thus, it was identified as Exiguobacterium acetylicum FHBD (accession number MW165834). Strain FHBD fermentation medium exhibited the maximum keratinase activity (2934.54 ± 38.56 U/mL) and sulfhydryl group content (3.09 ± 0.02 mM) at 72 h under optimal process conditions of pH 5.0, temperature (35 °C), inoculum size (2% v/v), and feather (15 g/L). Amino acid analysis of the feather hydrolysate showed significant levels of glutamic acid, aspartic acid, glycine, arginine, serine, and proline, with respective concentrations of 1.58, 1.34, 1.29, 1.20, 1.12, and 0.93 (g/100 g of sample). The study's findings emphasize the potential of E. acetylicum FHBD in poultry feather valorization and keratinase production.
Citation: Tutuka Dlume, Nonso E. Nnolim, Uchechukwu U. Nwodo. Exiguobacterium acetylicum transformed poultry feathers into amino acids through an extracellular secretion of keratinolytic enzymes[J]. AIMS Bioengineering, 2024, 11(4): 489-505. doi: 10.3934/bioeng.2024022
The transition from a traditional linear economy to a circular model aims to create a more sustainable future by reducing the adverse effects of agro-waste on the environment. The present study evaluated the metabolic diversity of bacterial isolates from municipal dumpsites for keratinase production and poultry feather valorization. The bacterium with the highest keratinolytic activity was identified through 16S ribosomal ribonucleic acid (rRNA) gene sequencing. The exo-keratinase production by the bacterium was optimized, and the feather hydrolysate obtained from the fermentation process was analyzed for amino acids. Among the twelve bacteria isolated from the dumpsite's sample, three showed significant feather degradation and keratinase production of 89% (744.5 ± 9.19 U/mL), 58% (269 ± 15.55 U/mL), and 46% (195 ± 7.07 U/mL) for SSB-03, SSB-02, and HSB-02, respectively. Analysis of the 16S rRNA gene sequence revealed that SSB-03 has high sequence homology with Exiguobacterium acetylicum, and thus, it was identified as Exiguobacterium acetylicum FHBD (accession number MW165834). Strain FHBD fermentation medium exhibited the maximum keratinase activity (2934.54 ± 38.56 U/mL) and sulfhydryl group content (3.09 ± 0.02 mM) at 72 h under optimal process conditions of pH 5.0, temperature (35 °C), inoculum size (2% v/v), and feather (15 g/L). Amino acid analysis of the feather hydrolysate showed significant levels of glutamic acid, aspartic acid, glycine, arginine, serine, and proline, with respective concentrations of 1.58, 1.34, 1.29, 1.20, 1.12, and 0.93 (g/100 g of sample). The study's findings emphasize the potential of E. acetylicum FHBD in poultry feather valorization and keratinase production.
Minimal salt media
Ground chicken feather
Polymerase chain reaction
National Centre for Biotechnology Information
Basic local alignment search tool

| [1] |
Muscat A, de Olde EM, Ripoll-Bosch R, et al. (2021) Principles, drivers and opportunities of a circular bioeconomy. Nat Food 2: 561-566. https://doi.org/10.1038/s43016-021-00340-7 
|
| [2] |
Velenturf AP, Purnell P (2021) Principles for a sustainable circular economy. Sustain Prod Consum 27: 1437-1457. https://doi.org/10.1016/j.spc.2021.02.018
|
| [3] |
Choi J, Kong B, Bowker BC, et al. (2023) Nutritional strategies to improve meat quality and composition in the challenging conditions of broiler production: a review. Animals 13: 1386. https://doi.org/10.3390/ani13081386
|
| [4] |
Zhang F, Jiang L, Wang S (2018) Repairable cascaded slide-lock system endows bird feathers with tear-resistance and superdurability. Proc Natl Acad Sci 115: 10046-10051. https://doi.org/10.1073/pnas.1808293115
|
| [5] |
Li ZW, Liang S, Ke Y, et al. (2020) The feather degradation mechanisms of a new Streptomyces sp. isolate SCUT-3. Commun Biol 3: 191. https://doi.org/10.1038/s42003-020-0918-0
|
| [6] |
Chilakamarry CR, Mahmood S, Saffe SNBM, et al. (2021) Extraction and application of keratin from natural resources: a review. 3 Biotech 11: 1-12. https://doi.org/10.1007/s13205-021-02734-7
|
| [7] |
Srivastava B, Khatri M, Singh G, et al. (2020) Microbial keratinases: an overview of biochemical characterization and its eco-friendly approach for industrial applications. J Clean Prod 252: 119847. https://doi.org/10.1016/j.jclepro.2019.119847
|
| [8] |
Dawan J, Ahn J (2022) Bacterial stress responses as potential targets in overcoming antibiotic resistance. Microorganisms 10: 1385. https://doi.org/10.3390/microorganisms10071385
|
| [9] |
Alwakeel SS, Ameen F, Al Gwaiz H, et al. (2021) Keratinases produced by Aspergillus stelliformis, Aspergillus sydowii, and Fusarium brachygibbosum isolated from human hair: yield and activity. J Fungi 7: 471. https://doi.org/10.3390/jof7060471
|
| [10] | Derhab N, Mabrouk ME, El-Metwally MM, et al. (2023) Thermostable keratinase from Bacillus cereus L10: optimization and some potential biotechnological applications. Biomass Convers Bioref 2023: 1-17. https://doi.org/10.1007/s13399-023-04887-2 |
| [11] |
Williams CM, Richter CS, MacKenzie Jr JM, et al. (1990) Isolation, identification, and characterization of a feather-degrading bacterium. Appl Environ Microbiol 56: 1509-1515. https://doi.org/10.1128/aem.56.6.1509-1515.1990
|
| [12] |
Yahaya RSR, Phang LY, Normi YM, et al. (2022) Feather-degrading Bacillus cereus HD1: genomic analysis and its optimization for keratinase production and feather degradation. Curr Microbiol 79: 166. https://doi.org/10.1007/s00284-022-02861-1
|
| [13] |
Peng Z, Mao X, Zhang J, et al. (2019) Effective biodegradation of chicken feather waste by co-cultivation of keratinase producing strains. Microb Cell Fact 18: 1-11. https://doi.org/10.1186/s12934-019-1134-9
|
| [14] |
Park G, Lee KM, Lee YS, et al. (2023) Biodegradation and valorization of feather waste using the keratinase-producing bacteria and their application in environmentally hazardous industrial processes. J Environ Manag 346: 118986. https://doi.org/10.1016/j.jenvman.2023.118986
|
| [15] |
Gurav RG, Tang J, Jadhav JP (2016) Sulfitolytic and keratinolytic potential of Chryseobacterium sp. RBT revealed hydrolysis of melanin containing feathers. 3 Biotech 6: 145. https://doi.org/10.1007/s13205-016-0464-0
|
| [16] |
Łaba W, Żarowska B, Chorążyk D, et al. (2018) New keratinolytic bacteria in valorization of chicken feather waste. AMB Express 8: 9. https://doi.org/10.1186/s13568-018-0538-y
|
| [17] |
Abdelmoteleb A, Gonzalez-Mendoza D, Tzintzun-Camacho O, et al. (2023) Keratinases from Streptomyces netropsis and Bacillus subtilis and their potential use in the chicken feather degrading. Fermentation 9: 96. https://doi.org/10.3390/fermentation9020096
|
| [18] |
Kshetri P, Singh PL, Chanu SB, et al. (2022) Biological activity of peptides isolated from feather keratin waste through microbial and enzymatic hydrolysis. Electron J Biotechnol 60: 11-18. https://doi.org/10.1016/j.ejbt.2022.08.001
|
| [19] | Lai Y, Wu X, Zheng X, et al. (2023) Insights into the keratin efficient degradation mechanism mediated by Bacillus sp. CN2 based on integrating functional degradomics. Biotechnol Biof Biop 16: 59. https://doi.org/10.1186/s13068-023-02308-0 |
| [20] |
El-Gindy AA, Ibrahim ZM, Aziz HM, et al. (2023) Chicken feather waste degradation by Malbranchea cinnamomea and its application on plant growth and metabolites of Vicia faba plant. Biocatal Agric Biotechnol 53: 102883. https://doi.org/10.1016/j.bcab.2023.102883
|
| [21] |
Zengler K, Zaramela LS (2018) The social network of microorganisms—how auxotrophies shape complex communities. Nat Rev Microbiol 16: 383-390. https://doi.org/10.1038/s41579-018-0004-5
|
| [22] |
Letourneau F, Soussotte V, Bressollier P, et al. (1998) Keratinolytic activity of Streptomyces sp. S. K1–02: a new isolated strain. Lett Appl Microbiol 26: 77-80. https://doi.org/10.1046/j.1472-765X.1998.00281.x
|
| [23] |
Riffel A, Brandelli A (2006) Keratinolytic bacteria isolated from feather waste. Braz J Microbiol 37: 395-399. https://doi.org/10.1590/S1517-83822006000300036
|
| [24] |
Bokveld A, Nnolim NE, Digban TO, et al. (2023) Chryseobacterium aquifrigidense keratinase liberated essential and nonessential amino acids from chicken feather degradation. Environ Technol 44: 293-303. https://doi.org/10.1080/09593330.2021.1969597
|
| [25] |
Nnolim NE, Okoh AI, Nwodo UU (2020) Proteolytic bacteria isolated from agro-waste dumpsites produced keratinolytic enzymes. Biotechnol Rep 27: e00483. https://doi.org/10.1016/j.btre.2020.e00483
|
| [26] |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254. https://doi.org/10.1016/0003-2697(76)90527-3
|
| [27] |
Ellman GL (1959) Tissue sulfhydryl groups. Arch Biochem Biophys 82: 70-77. https://doi.org/10.1016/0003-9861(59)90090-6
|
| [28] |
Turner S, Pryer KM, Miao VP, et al. (1999) Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis1. J Eukaryot Microbiol 46: 327-338. https://doi.org/10.1111/j.1550-7408.1999.tb04612.x
|
| [29] |
Altschul SF, Madden TL, Schäffer AA, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389-3402. https://doi.org/10.1093/nar/25.17.3389
|
| [30] |
Einarsson S, Josefsson B, Lagerkvist S (1983) Determination of amino acids with 9-fluorenylmethyl chloroformate and reversed-phase high-performance liquid chromatography. J Chromatogr A 282: 609-618. https://doi.org/10.1016/S0021-9673(00)91638-8
|
| [31] |
Bank RA, Jansen EJ, Beekman B, et al. (1996) Amino acid analysis by reverse-phase high-performance liquid chromatography: improved derivatization and detection conditions with 9-fluorenylmethyl chloroformate. Anal Biochem 240: 167-176. https://doi.org/10.1006/abio.1996.0346
|
| [32] |
Ke H, Li J, Zhang X, et al. (2022) Bacterial community structure and predicted metabolic function of landfilled municipal solid waste in China. Sustainability 14: 3144. https://doi.org/10.3390/su14063144
|
| [33] | Gumilar J, Triatmojo S, Yusiati LM, et al. (2015) Isolation, identification and dehairing activity of Indonesian native keratinolytic bacteria Exiguobacterium sp. DG1. Pak J Biotechnol 12: 41-48. https://pjbt.org/index.php/pjbt/article/view/128 |
| [34] |
Emon TH, Hakim A, Chakraborthy D, et al. (2020) Kinetics, detergent compatibility and feather-degrading capability of alkaline protease from Bacillus subtilis AKAL7 and Exiguobacterium indicum AKAL11 produced with fermentation of organic municipal solid wastes. J Environ Sci Health Part A 55: 1339-1348. https://doi.org/10.1080/10934529.2020.1794207
|
| [35] |
Alster CJ, Baas P, Wallenstein MD, et al. (2016) Temperature sensitivity as a microbial trait using parameters from macromolecular rate theory. Front Microbiol 7: 1821. https://doi.org/10.3389/fmicb.2016.01821
|
| [36] |
Revankar AG, Bagewadi ZK, Bochageri NP, et al. (2023) Response surface methodology based optimization of keratinase from Bacillus velezensis strain ZBE1 and nanoparticle synthesis, biological and molecular characterization. Saudi J Biol Sci 30: 103787. https://doi.org/10.1016/j.sjbs.2023.103787
|
| [37] |
Jin Q, Kirk MF (2018) pH as a primary control in environmental microbiology: 1. thermodynamic perspective. Front Environ Sci 6: 21. https://doi.org/10.3389/fenvs.2018.00021
|
| [38] |
Jiang J, Sun YF, Tang X, et al. (2018) Alkaline pH shock enhanced production of validamycin a in fermentation of Streptomyces hygroscopicus. Bioresour Technol 249: 234-240. https://doi.org/10.1016/j.biortech.2017.10.012
|
| [39] |
Kim JM, Lim WJ, Suh HJ (2001) Feather-degrading bacillus species from poultry waste. Process Biochem 37: 287-291. https://doi.org/10.1016/S0032-9592(01)00206-0
|
| [40] |
Nnolim NE, Okoh AI, Nwodo UU (2020b) Bacillus sp. FPF-1 produced keratinase with high potential for chicken feather degradation. Molecules 25: 1505. https://doi.org/10.3390/molecules25071505
|
| [41] |
Abusham RA, Rahman RNZR, Salleh AB, et al. (2009) Optimization of physical factors affecting the production of thermo-stable organic solvent-tolerant protease from a newly isolated halo tolerant Bacillus subtilis strain rand. Microb Cell Factories 8: 20. https://doi.org/10.1186/1475-2859-8-20
|
| [42] |
Cai CG, Lou BG, Zheng XD (2008) Keratinase production and keratin degradation by a mutant strain of Bacillus subtilis. J Zhejiang Univ Sci B 9: 60-67. https://doi.org/10.1631/jzus.B061620
|
| [43] |
Parashar D, Bhatia D, Malik DK (2017) Optimization of keratinase production by Bacillus olironius isolated from poultry farm soil. J Pure Appl Microbiol 11: 1129-1134. https://doi.org/10.22207/JPAM.11.2.58
|
| [44] |
Barman NC, Zohora FT, Das KC, et al. (2017) Production, partial optimization and characterization of keratinase enzyme by Arthrobacter sp. NFH5 isolated from soil samples. AMB Express 7: 181. https://doi.org/10.1186/s13568-017-0462-6
|
| [45] | Zhang YZ, Zhang WX, Chen XL (2020) Mechanisms for induction of microbial extracellular proteases in response to exterior proteins. Appl Environ Microbiol 86: e01036-20. https://doi.org/10.1128/AEM.01036-20 |
| [46] |
Shen N, Yang M, Xie C, et al. (2022) Isolation and identification of a feather degrading Bacillus tropicus strain gxun-17 from marine environment and its enzyme characteristics. BMC Biotechnol 22: 11. https://doi.org/10.1186/s12896-022-00742-w
|
| [47] |
Wang L, Qian Y, Cao Y, et al. (2017) Production and characterization of keratinolytic proteases by a chicken feather-degrading thermophilic strain, Thermoactinomyces sp. YT06. J Microbiol Biotechnol 27: 2190-2198. https://doi.org/10.4014/jmb.1705.05082
|
| [48] | Wu XQ, Chen L, Cao ZJ, et al. (2012) Feather degradation and keratinase production by Stenotrophomonas maltophilia DHHJ. Adv Mat Res 550: 1400-1403. https://doi.org/10.4028/www.scientific.net/AMR.550-553.1400 |
| [49] |
Jaishankar J, Srivastava P (2017) Molecular basis of stationary phase survival and applications. Front Microbiol 8: 2000. https://doi.org/10.3389/fmicb.2017.02000
|
| [50] |
Pletnev P, Osterman I, Sergiev P, et al. (2015) Survival guide: Escherichia coli in the stationary phase. Acta Nature 7: 22-33. https://doi.org/10.32607/20758251-2015-7-4-22-33
|
| [51] |
Yamamura S, Morita Y, Hasan Q, et al. (2002) Keratin degradation: a cooperative action of two enzymes from Stenotrophomonas sp. Biochem Biophys Res Commun 294: 1138-1143. https://doi.org/10.1016/S0006-291X(02)00580-6
|
| [52] |
Jeong JH, Park KH, Oh DJ, et al. (2010) Keratinolytic enzyme-mediated biodegradation of recalcitrant feather by a newly isolated Xanthomonas sp. P5. Polym Degrad Stab 95: 1969-1977. https://doi.org/10.1016/j.polymdegradstab.2010.07.020
|
| [53] |
Peng S, Li H, Zhang S, et al. (2023) Isolation of a novel feather-degrading Ectobacillus sp. JY-23 strain and characterization of a new keratinase in the M4 metalloprotease family. Microbiol Res 274: 127439. https://doi.org/10.1016/j.micres.2023.127439
|
| [54] |
Sandström V, Chrysafi A, Lamminen M, et al. (2022) Food system by-products upcycled in livestock and aquaculture feeds can increase global food supply. Nat Food 3: 729-740. https://doi.org/10.1038/s43016-022-00589-6
|
| [55] |
Callegaro K, Brandelli A, Daroit DJ (2019) Beyond plucking: feathers bioprocessing into valuable protein hydrolysates. Waste Manag 95: 399-415. https://doi.org/10.1016/j.wasman.2019.06.040
|
| [56] |
Zhang J, Su C, Kong XL, et al. (2022) Directed evolution driving the generation of an efficient keratinase variant to facilitate the feather degradation. Bioresour Bioprocess 9: 38. https://doi.org/10.1186/s40643-022-00524-4
|
| [57] |
Safari H, Mohit A, Mohiti-Asli M (2024) Feather meal processing methods impact the production parameters, blood biochemical indices, gut function, and hepatic enzyme activity in broilers. J Anim Sci 102: skae068. https://doi.org/10.1093/jas/skae068
|
| [58] | Fitriyanto NA, Ramadhanti Y, Rusyadi I, et al. (2022) Production of poultry feather hydrolysate using HCl and NaOH as a growth medium substrate for indigenous strains. Earth Environ Sci 951: 012064. https://doi.org/10.1088/1755-1315/951/1/012064 |
| [59] |
Tamreihao K, Devi LJ, Khunjamayum R, et al. (2017) Biofertilizing potential of feather hydrolysate produced by indigenous keratinolytic Amycolatopsis sp. MBRL 40 for rice cultivation under field conditions. Biocatal Agri Biotechnol 10: 317-320. https://doi.org/10.1016/j.bcab.2017.04.010
|
| [60] |
El Salamony DH, Hassouna MSE, Zaghloul TI, et al. (2024) Bioenergy production from chicken feather waste by anaerobic digestion and bioelectrochemical systems. Microb Cell Fact 23: 102. https://doi.org/10.1186/s12934-024-02374-5
|

Figures(5)

Tutuka Dlume, Nonso E. Nnolim, Uchechukwu U. Nwodo. Exiguobacterium acetylicum transformed poultry feathers into amino acids through an extracellular secretion of keratinolytic enzymes[J]. AIMS Bioengineering, 2024, 11(4): 489-505. doi: 10.3934/bioeng.2024022







 DownLoad:
DownLoad: