Citation: F. Philipp Seib. Silk nanoparticles—an emerging anticancer nanomedicine[J]. AIMS Bioengineering, 2017, 4(2): 239-258. doi: 10.3934/bioeng.2017.2.239

| [1] |
Vollrath F, Porter D (2009) Silks as ancient models for modern polymers. Polymer 50: 5623–5632. doi: 10.1016/j.polymer.2009.09.068

|
| [2] | Lubec G, Holaubek J, Feldl C, et al. (1993) Use of silk in ancient Egypt. Nature 362: 25. |
| [3] |
Altman GH, Diaz F, Jakuba C, et al. (2003) Silk-based biomaterials. Biomaterials 24: 401–416. doi: 10.1016/S0142-9612(02)00353-8
|
| [4] |
Omenetto FG, Kaplan DL (2010) New opportunities for an ancient material. Science 329: 528–531. doi: 10.1126/science.1188936
|
| [5] |
Elices M, Plaza GR, Perez RJ, et al. (2011) The hidden link between supercontraction and mechanical behavior of spider silks. J Mech Behav Biomed Mater 4: 658–669. doi: 10.1016/j.jmbbm.2010.09.008
|
| [6] |
Cranford SW, Tarakanova A, Pugno NM, et al. (2012) Nonlinear material behaviour of spider silk yields robust webs. Nature 482: 72–76. doi: 10.1038/nature10739
|
| [7] |
Vollrath F, Porter D, Holland C (2013) The science of silks. MRS Bull 38: 73–80. doi: 10.1557/mrs.2012.314
|
| [8] |
Kluge JA, Rabotyagova O, Leisk GG, et al. (2008) Spider silks and their applications. Trends Biotechnol 26: 244–251. doi: 10.1016/j.tibtech.2008.02.006
|
| [9] |
Gatesy J, Hayashi C, Motriuk D, et al. (2001) Extreme diversity, conservation, and convergence of spider silk fibroin sequences. Science 291: 2603–2605. doi: 10.1126/science.1057561
|
| [10] |
Hardy JG, Scheibel TR (2009) Silk-inspired polymers and proteins. Biochem Soc T 37: 677–681. doi: 10.1042/BST0370677
|
| [11] |
Rising A, Johansson J (2015) Toward spinning artificial spider silk. Nat Chem Biol 11: 309–315. doi: 10.1038/nchembio.1789
|
| [12] |
Kim S, Marelli B, Brenckle MA, et al. (2014) All-water-based electron-beam lithography using silk as a resist. Nat Nanotechnol 9: 306–310. doi: 10.1038/nnano.2014.47
|
| [13] |
Omenetto FG, Kaplan DL (2008) A new route for silk. Nature Photonic 2: 641–643. doi: 10.1038/nphoton.2008.207
|
| [14] | Zhu B, Wang H, Leow WR, et al. (2016) Silk fibroin for flexible electronic devices. Adv Mater 22: 4250–4265. |
| [15] |
Doblhofer E, Schmid J, Riess M, et al. (2016) Structural insights into water-based spider silk protein-nanoclay composites with excellent gas and water vapor barrier properties. ACS Appl Mater Interface 8: 25535–25543. doi: 10.1021/acsami.6b08287
|
| [16] |
Marelli B, Brenckle MA, Kaplan DL, et al. (2016) Silk fibroin as edible coating for perishable food preservation. Sci Rep 6: 25263–25273. doi: 10.1038/srep25263
|
| [17] |
Abbott RD, Kimmerling EP, Cairns DM, et al. (2016) Silk as a biomaterial to support long-term three-dimensional tissue cultures. ACS Appl Mater Interface 8: 21861–21868. doi: 10.1021/acsami.5b12114
|
| [18] |
Jao D, Mou X, Hu X (2016) Tissue regeneration: a silk road. J Funct Biomater 7: 22–39. doi: 10.3390/jfb7030022
|
| [19] |
Kasoju N, Bora U (2012) Silk fibroin in tissue engineering. Adv Healthc Mater 1: 393–412. doi: 10.1002/adhm.201200097
|
| [20] |
Werner V, Meinel L (2015) From silk spinning in insects and spiders to advanced silk fibroin drug delivery systems. Eur J Pharm Biopharm 97: 392–399. doi: 10.1016/j.ejpb.2015.03.016
|
| [21] |
Yucel T, Lovett ML, Kaplan DL (2014) Silk-based biomaterials for sustained drug delivery. J Control Release 190: 381–397. doi: 10.1016/j.jconrel.2014.05.059
|
| [22] | Seib FP, Kaplan DL (2013) Silk for drug delivery applications: opportunities and challenges. Israel J Chem 53: 756–766. |
| [23] |
Thurber AE, Omenetto FG, Kaplan DL (2015) In vivo bioresponses to silk proteins. Biomaterials 71: 145–157. doi: 10.1016/j.biomaterials.2015.08.039
|
| [24] |
Pritchard EM, Dennis PB, Omenetto FG, et al. (2012) Physical and chemical aspects of stabilization of compounds in silk. Biopolymers 97: 479–498. doi: 10.1002/bip.22026
|
| [25] |
Chiu B, Coburn J, Pilichowska M, et al. (2014) Surgery combined with controlled-release doxorubicin silk films as a treatment strategy in an orthotopic neuroblastoma mouse model. Brit J Cancer 111: 708–715. doi: 10.1038/bjc.2014.324
|
| [26] |
Seib FP, Coburn J, Konrad I, et al. (2015) Focal therapy of neuroblastoma using silk films to deliver kinase and chemotherapeutic agents in vivo. Acta Biomater 20: 32–38. doi: 10.1016/j.actbio.2015.04.003
|
| [27] |
Coburn J, Harris J, Zakharov AD, et al. (2017) Implantable chemotherapy-loaded silk protein materials for neuroblastoma treatment. Int J Cancer 140: 726–735. doi: 10.1002/ijc.30479
|
| [28] |
Seib FP, Pritchard EM, Kaplan DL (2013) Self-assembling doxorubicin silk hydrogels for the focal treatment of primary breast cancer. Adv Funct Mater 23: 58–65. doi: 10.1002/adfm.201201238
|
| [29] |
Jastrzebska K, Kucharczyk K, Florczak A, et al. (2015) Silk as an innovative biomaterial for cancer therapy. Rep Pract Oncol Radiother 20: 87–98. doi: 10.1016/j.rpor.2014.11.010
|
| [30] | Coleman RE (2012) Bone cancer in 2011: prevention and treatment of bone metastases. Nat Rev Clin Oncol 9: 76–78. |
| [31] |
Gupta GP, Massague J (2006) Cancer metastasis: building a framework. Cell 127: 679–695. doi: 10.1016/j.cell.2006.11.001
|
| [32] |
Ehrlich P (1913) Address in pathology, on chemiotherapy: delivered before the seventeenth international congress of medicine. Brit Med J 2: 353–359. doi: 10.1136/bmj.2.2746.353
|
| [33] |
Mottaghitalab F, Farokhi M, Shokrgozar MA, et al. (2015) Silk fibroin nanoparticle as a novel drug delivery system. J Control Release 206: 161–176. doi: 10.1016/j.jconrel.2015.03.020
|
| [34] |
Zhao Z, Li Y, Xie MB (2015) Silk fibroin-based nanoparticles for drug delivery. Int J Mol Sci 16: 4880–4903. doi: 10.3390/ijms16034880
|
| [35] |
Ebrahimi D, Tokareva O, Rim NG, et al. (2015) Silk-its mysteries, how it is made, and how it is used. ACS Biomater Sci Eng 1: 864–876. doi: 10.1021/acsbiomaterials.5b00152
|
| [36] |
Eisoldt L, Thamm C, Scheibel T (2012) Review the role of terminal domains during storage and assembly of spider silk proteins. Biopolymers 97: 355–361. doi: 10.1002/bip.22006
|
| [37] | Xu G, Gong L, Yang Z, et al. (2014) What makes spider silk fibers so strong? From molecular-crystallite network to hierarchical network structures. Soft Mat 10: 2116–2123. |
| [38] |
Ha SW, Gracz HS, Tonelli AE, et al. (2005) Structural study of irregular amino acid sequences in the heavy chain of bombyx mori silk fibroin. Biomacromolecules 6: 2563–2569. doi: 10.1021/bm050294m
|
| [39] |
Asakura T, Ohgo K, Ishida T, et al. (2005) Possible implications of serine and tyrosine residues and intermolecular interactions on the appearance of silk istructure of bombyx mori silk fibroin-derived synthetic peptides: high-resolution 13c cross-polarization/magic-angle spinning NMR study. Biomacromolecules 6: 468–474. doi: 10.1021/bm049487k
|
| [40] |
Asakura T, Okushita K, Williamson MP (2015) Analysis of the structure of bombyx mori silk fibroin by NMR. Macromolecules 48: 2345–2357. doi: 10.1021/acs.macromol.5b00160
|
| [41] |
Zhou CZ, Confalonieri F, Jacquet M, et al. (2001) Silk fibroin: structural implications of a remarkable amino acid sequence. Proteins 44: 119–122. doi: 10.1002/prot.1078
|
| [42] |
Zhou CZ, Confalonieri F, Medina N, et al. (2000) Fine organization of bombyx mori fibroin heavy chain gene. Nucleic Acids Res 28: 2413–2419. doi: 10.1093/nar/28.12.2413
|
| [43] |
Jin HJ, Kaplan DL (2003) Mechanism of silk processing in insects and spiders. Nature 424: 1057–1061. doi: 10.1038/nature01809
|
| [44] |
Greving I, Dicko C, Terry A, et al. (2010) Small angle neutron scattering of native and reconstituted silk fibroin. Soft Mat 6: 4389–4395. doi: 10.1039/c0sm00108b
|
| [45] |
Lu Q, Zhu H, Zhang C, et al. (2012) Silk self-assembly mechanisms and control from thermodynamics to kinetics. Biomacromolecules 13: 826–832. doi: 10.1021/bm201731e
|
| [46] |
Wang X, Yucel T, Lu Q, et al. (2010) Silk nanospheres and microspheres from silk/pva blend films for drug delivery. Biomaterials 31: 1025–1035. doi: 10.1016/j.biomaterials.2009.11.002
|
| [47] |
Myung SJ, Kim HS, Kim Y, et al. (2008) Fluorescent silk fibroin nanoparticles prepared using a reverse microemulsion. Macromol Res 16: 604–608. doi: 10.1007/BF03218567
|
| [48] | Gupta V, Aseh A, Rios CN, et al. (2009) Fabrication and characterization of silk fibroin-derived curcumin nanoparticles for cancer therapy. Int J Nanomed 4: 115–122. |
| [49] |
Lammel AS, Hu X, Park SH, et al. (2010) Controlling silk fibroin particle features for drug delivery. Biomaterials 31: 4583–4591. doi: 10.1016/j.biomaterials.2010.02.024
|
| [50] |
Kundu J, Chung YI, Kim YH, et al. (2010) Silk fibroin nanoparticles for cellular uptake and control release. Int J Pharm 388: 242–250. doi: 10.1016/j.ijpharm.2009.12.052
|
| [51] |
Seib FP, Jones GT, Rnjak KJ, et al. (2013) pH-dependent anticancer drug release from silk nanoparticles. Adv Healthc Mater 2: 1606–1611. doi: 10.1002/adhm.201300034
|
| [52] | Wongpinyochit T, Johnston BF, Seib FP (2016) Manufacture and drug delivery applications of silk nanoparticles. J Vis Exp DOI: 10.3791/54669. |
| [53] |
Zhang YQ, Shen WD, Xiang RL, et al. (2007) Formation of silk fibroin nanoparticles in water-miscible organic solvent and their characterization. J Nanopart Res 9: 885–900. doi: 10.1007/s11051-006-9162-x
|
| [54] | Zhao Z, Xie M, Li Y, et al. (2015) Formation of curcumin nanoparticles via solution-enhanced dispersion by supercritical CO2. Int J Nanomed 10: 3171–3181. |
| [55] | Lozano PAA, Montalban MG, Aznar CSD, et al. (2015) Production of silk fibroin nanoparticles using ionic liquids and high-power ultrasounds. J Appl Polym Sci 132: 41702–41709. |
| [56] | Gholami A, Tavanai H, Moradi AR (2010) Production of fibroin nanopowder through electrospraying. J Nanopart Res 13: 2089–2098. |
| [57] |
Wenk E, Wandrey AJ, Merkle HP, et al. (2008) Silk fibroin spheres as a platform for controlled drug delivery. J Control Release 132: 26–34. doi: 10.1016/j.jconrel.2008.08.005
|
| [58] |
Lu Q, Huang Y, Li M, et al. (2011) Silk fibroin electrogelation mechanisms. Acta Biomater 7: 2394–2400. doi: 10.1016/j.actbio.2011.02.032
|
| [59] |
Rajkhowa R, Wang L, Wang X (2008) Ultra-fine silk powder preparation through rotary and ball milling. Powder Technol 185: 87–95. doi: 10.1016/j.powtec.2008.01.005
|
| [60] |
Mathur AB, Gupta V (2010) Silk fibroin-derived nanoparticles for biomedical applications. Nanomedicine 5: 807–820. doi: 10.2217/nnm.10.51
|
| [61] |
Xiao L, Lu G, Lu Q, et al. (2016) Direct formation of silk nanoparticles for drug delivery. ACS Biomater Sci Eng 2: 2050–2057. doi: 10.1021/acsbiomaterials.6b00457
|
| [62] |
Wongpinyochit T, Uhlmann P, Urquhart AJ, et al. (2015) PEGylated silk nanoparticles for anticancer drug delivery. Biomacromolecules 16: 3712–3722. doi: 10.1021/acs.biomac.5b01003
|
| [63] |
Subia B, Chandra S, Talukdar S, et al. (2014) Folate conjugated silk fibroin nanocarriers for targeted drug delivery. Integr Biol 6: 203–214. doi: 10.1039/C3IB40184G
|
| [64] |
Rabanel JM, Hildgen P, Banquy X (2014) Assessment of PEG on polymeric particles surface, a key step in drug carrier translation. J Control Release 185: 71–87. doi: 10.1016/j.jconrel.2014.04.017
|
| [65] |
Pasut G, Veronese FM (2012) State of the art in PEGylation: the great versatility achieved after forty years of research. J Control Release 161: 461–472. doi: 10.1016/j.jconrel.2011.10.037
|
| [66] |
Wang S, Xu T, Yang Y, et al. (2015) Colloidal stability of silk fibroin nanoparticles coated with cationic polymer for effective drug delivery. ACS Appl Mater Interface 7: 21254–21262. doi: 10.1021/acsami.5b05335
|
| [67] |
Tian Y, Jiang X, Chen X, et al. (2014) Doxorubicin-loaded magnetic silk fibroin nanoparticles for targeted therapy of multidrug-resistant cancer. Adv Mater 26: 7393–7398. doi: 10.1002/adma.201403562
|
| [68] |
Chung H, Kim TY, Lee SY (2012) Recent advances in production of recombinant spider silk proteins. Curr Opin Biotech 23: 957–964. doi: 10.1016/j.copbio.2012.03.013
|
| [69] |
Lammel A, Schwab M, Hofer M, et al. (2011) Recombinant spider silk particles as drug delivery vehicles. Biomaterials 32: 2233–2240. doi: 10.1016/j.biomaterials.2010.11.060
|
| [70] |
Humenik M, Smith AM, Scheibel T (2011) Recombinant spider silks-biopolymers with potential for future applications. Polymers 3: 640–661. doi: 10.3390/polym3010640
|
| [71] |
Schierling MB, Doblhofer E, Scheibel T (2016) Cellular uptake of drug loaded spider silk particles. Biomater Sci 4: 1515–1523. doi: 10.1039/C6BM00435K
|
| [72] |
Doblhofer E, Scheibel T (2015) Engineering of recombinant spider silk proteins allows defined uptake and release of substances. J Pharm Sci 104: 988–994. doi: 10.1002/jps.24300
|
| [73] |
Elsner MB, Herold HM, Muller HS, et al. (2015) Enhanced cellular uptake of engineered spider silk particles. Biomater Sci 3: 543–551. doi: 10.1039/C4BM00401A
|
| [74] |
Florczak A, Mackiewicz A, Dams KH (2014) Functionalized spider silk spheres as drug carriers for targeted cancer therapy. Biomacromolecules 15: 2971–2981. doi: 10.1021/bm500591p
|
| [75] |
Neubauer MP, Blüm C, Agostini E, et al. (2013) Micromechanical characterization of spider silk particles. Biomater Sci 1: 1160–1165. doi: 10.1039/c3bm60108k
|
| [76] |
Anselmo AC, Zhang M, Kumar S, et al. (2015) Elasticity of nanoparticles influences their blood circulation, phagocytosis, endocytosis, and targeting. ACS Nano 9: 3169–3177. doi: 10.1021/acsnano.5b00147
|
| [77] |
Numata K, Kaplan DL (2010) Silk-based delivery systems of bioactive molecules. Adv Drug Deliver Rev 62: 1497–1508. doi: 10.1016/j.addr.2010.03.009
|
| [78] |
Numata K, Subramanian B, Currie HA, et al. (2009) Bioengineered silk protein-based gene delivery systems. Biomaterials 30: 5775–5784. doi: 10.1016/j.biomaterials.2009.06.028
|
| [79] |
Numata K, Hamasaki J, Subramanian B, et al. (2010) Gene delivery mediated by recombinant silk proteins containing cationic and cell binding motifs. J Control Release 146: 136–143. doi: 10.1016/j.jconrel.2010.05.006
|
| [80] |
Numata K, Kaplan DL (2010) Silk-based gene carriers with cell membrane destabilizing peptides. Biomacromolecules 11: 3189–3195. doi: 10.1021/bm101055m
|
| [81] |
Numata K, Reagan MR, Goldstein RH, et al. (2011) Spider silk-based gene carriers for tumor cell-specific delivery. Bioconjugate Chem 22: 1605–1610. doi: 10.1021/bc200170u
|
| [82] |
Seib FP, Herklotz M, Burke KA, et al. (2014) Multifunctional silk-heparin biomaterials for vascular tissue engineering applications. Biomaterials 35: 83–91. doi: 10.1016/j.biomaterials.2013.09.053
|
| [83] |
Seib FP, Maitz MF, Hu X, et al. (2012) Impact of processing parameters on the haemocompatibility of bombyx mori silk films. Biomaterials 33: 1017–1023. doi: 10.1016/j.biomaterials.2011.10.063
|
| [84] |
Murphy AR, Kaplan DL (2009) Biomedical applications of chemically-modified silk fibroin. J Mater Chem 19: 6443–6450. doi: 10.1039/b905802h
|
| [85] |
Kambe Y, Yamamoto K, Kojima K, et al. (2010) Effects of RGDS sequence genetically interfused in the silk fibroin light chain protein on chondrocyte adhesion and cartilage synthesis. Biomaterials 31: 7503–7511. doi: 10.1016/j.biomaterials.2010.06.045
|
| [86] |
Teule F, Miao YG, Sohn BH, et al. (2012) Silkworms transformed with chimeric silkworm/spider silk genes spin composite silk fibers with improved mechanical properties. P Natl Acad Sci USA 109: 923–928. doi: 10.1073/pnas.1109420109
|
| [87] |
Xia XX, Qian ZG, Ki CS, et al. (2010) Native-sized recombinant spider silk protein produced in metabolically engineered Escherichia coli results in a strong fiber. P Natl Acad Sci USA 107: 14059–14063. doi: 10.1073/pnas.1003366107
|
| [88] | Wray LS, Hu X, Gallego J, et al. (2011) Effect of processing on silk-based biomaterials: reproducibility and biocompatibility. J Biomed Mater Res 99: 89–101. |
| [89] |
Rockwood DN, Preda RC, Yucel T, et al. (2011) Materials fabrication from bombyx mori silk fibroin. Nat Protoc 6: 1612–1631. doi: 10.1038/nprot.2011.379
|
| [90] |
Duncan R, Gaspar R (2011) Nanomedicine(s) under the microscope. Mol Pharm 8: 2101–2141. doi: 10.1021/mp200394t
|
| [91] |
Sheridan C (2012) Proof of concept for next-generation nanoparticle drugs in humans. Nature Biotechnol 30: 471–473. doi: 10.1038/nbt0612-471
|
| [92] | Shi J, Kantoff PW, Wooster R, et al. (2017) Cancer nanomedicine: progress, challenges and opportunities. Nat Rev Cancer 17: 20–37. |
| [93] |
Maeda H, Nakamura H, Fang J (2013) The EPR effect for macromolecular drug delivery to solid tumors: improvement of tumor uptake, lowering of systemic toxicity, and distinct tumor imaging in vivo. Adv Drug Deliver Rev 65: 71–79. doi: 10.1016/j.addr.2012.10.002
|
| [94] |
Duncan R (2006) Polymer conjugates as anticancer nanomedicines. Nat Rev Cancer 6: 688–701. doi: 10.1038/nrc1958
|
| [95] |
Juliano R (2013) Nanomedicine: is the wave cresting? Nat Rev Drug Discov 12: 171–172. doi: 10.1038/nrd3958
|
| [96] | Venditto VJ, Szoka FC (2013) Cancer nanomedicines: so many papers and so few drugs! Adv Drug Deliver Rev 65: 80–88. |
| [97] | Wilhelm S, Tavares AJ, Dai Q, et al. (2016) Analysis of nanoparticle delivery to tumours. Nature Rev Mater 1: 1–12. |
| [98] |
Cleal K, He L, Watson PD, et al. (2013) Endocytosis, intracellular traffic and fate of cell penetrating peptide based conjugates and nanoparticles. Curr Pharm Design 19: 2878–2894. doi: 10.2174/13816128113199990297
|
| [99] |
Duncan R, Richardson SC (2012) Endocytosis and intracellular trafficking as gateways for nanomedicine delivery: opportunities and challenges. Mol Pharm 9: 2380–2402. doi: 10.1021/mp300293n
|
| [100] |
Whitehead KA, Langer R, Anderson DG (2009) Knocking down barriers: advances in siRNA delivery. Nat Rev Drug Discov 8: 129–138. doi: 10.1038/nrd2742
|
| [101] |
Gratton SE, Ropp PA, Pohlhaus PD, et al. (2008) The effect of particle design on cellular internalization pathways. P Natl Acad Sci USA 105: 11613–11618. doi: 10.1073/pnas.0801763105
|
| [102] |
Herd H, Daum N, Jones AT, et al. (2013) Nanoparticle geometry and surface orientation influence mode of cellular uptake. ACS Nano 7: 1961–1973. doi: 10.1021/nn304439f
|
| [103] |
Rejman J, Oberle V, Zuhorn IS, et al. (2004) Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem J 377: 159–169. doi: 10.1042/bj20031253
|
| [104] |
Oh P, Borgstrom P, Witkiewicz H, et al. (2007) Live dynamic imaging of caveolae pumping targeted antibody rapidly and specifically across endothelium in the lung. Nat Biotechnol 25: 327–337. doi: 10.1038/nbt1292
|
| [105] |
Sabharanjak S, Mayor S (2004) Folate receptor endocytosis and trafficking. Adv Drug Deliver Rev 56: 1099–1109. doi: 10.1016/j.addr.2004.01.010
|
| [106] |
Mosesson Y, Mills GB, Yarden Y (2008) Derailed endocytosis: an emerging feature of cancer. Nat Rev Cancer 8: 835–850. doi: 10.1038/nrc2521
|
| [107] |
Vercauteren D, Vandenbroucke RE, Jones AT, et al. (2010) The use of inhibitors to study endocytic pathways of gene carriers: optimization and pitfalls. Mol Ther 18: 561–569. doi: 10.1038/mt.2009.281
|
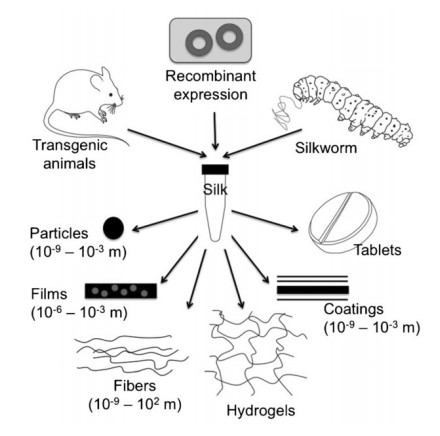
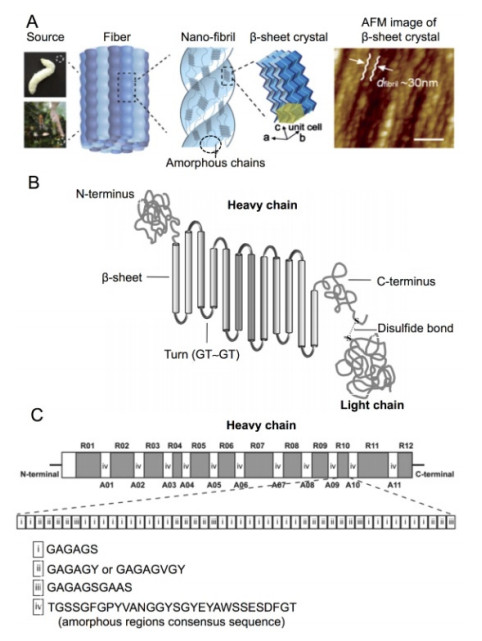
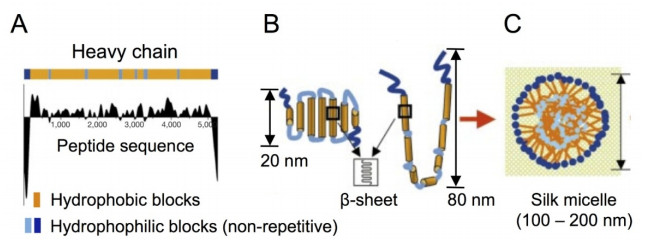
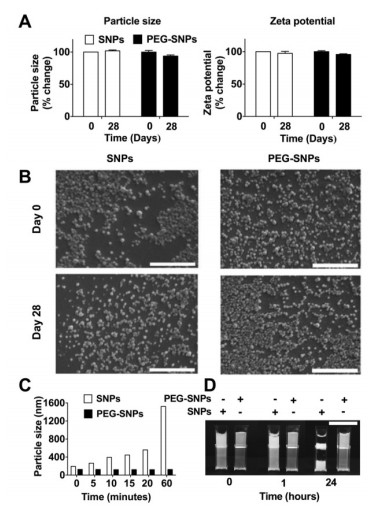
Figures(5)

F. Philipp Seib. Silk nanoparticles—an emerging anticancer nanomedicine[J]. AIMS Bioengineering, 2017, 4(2): 239-258. doi: 10.3934/bioeng.2017.2.239







 DownLoad:
DownLoad: