Citation: Lindsay Arnold, Rachel Chen. One-step non-chromatography purification of a low abundant fucosylated protein from complex plant crude extract[J]. AIMS Bioengineering, 2015, 2(3): 249-263. doi: 10.3934/bioeng.2015.3.249

| [1] | Ghazarian H, Idoni B, Oppenheimer SB. (2011) A glycobiology review: carbohydrates, lectins and implications in cancer therapeutics. Acta Histochem ,113: 236-247. |
| [2] | Wiederschain GY (2013) Glycobiology: progress, problems, and perspectives. Biochem Biokhimiia ,78: 679-696. |
| [3] | Varki A, Cummings RD, Esko JD, et al. (2009) Essentials of Glycobiology. 2nd ed. Cold Spring Harbor, editor. NY: Cold Spring Harbor Laboratory Press . |
| [4] | Ekins S, Xu JJ (2009) Drug Efficacy, Safety, and Biologics Discovery: Emerging Technologies and Tools. Hoboken, New Jersey: John Wiley & Sons, Inc . |
| [5] | Jefferis R (2009) Glycosylation as a strategy to improve antibody-based therapeutics. Nat Rev Drug Discov ,8: 226-234. |
| [6] | Nakano M, Kondo A, Kakehi K, et al. (2006) Glycomics - a new target for pharmaceuticals. Drug Discov Today Technol ,3: 39-47. |
| [7] | Raman R, Venkataraman M, Ramakrishnan S, et al. (2006) Advancing glycomics: implementation strategies at the consortium for functional glycomics. Glycobiology ,16: 82R-90R. |
| [8] | Taniguchi N, Ekuni A, Ko JH, et al. (2001) A glycomic approach to the identification and characterization of glycoprotein function in cells transfected with glycosyltransferase genes. Proteomics ,1: 239-247. |
| [9] | Gabius H-J, André S, Jiménez-Barbero J, et al. (2011) From lectin structure to functional glycomics: principles of the sugar code. Trends Biochem Sci ,36: 298-313. |
| [10] | VanHaandel MJ, Sarabèr FC, Boersma MG, et al. (2000) Characterization of different commercial soybean peroxidase preparations and use of the enzyme for N-demethylation of methyl N-methylanthranilate To produce the food flavor methylanthranilate. J Agric Food Chem ,48: 1949-1954. |
| [11] | Chattopadhyay K, Mazumdar S (2000) Structural and Conformational Stability of Horseradish Peroxidase: Effect of Temperature and pH. Biochemistry ,39: 263-270. |
| [12] | Sakharov IY, Berlina AN, Zherdev AV, et al. (2010) Advantages of soybean peroxidase over horseradish peroxidase as the enzyme label in chemiluminescent enzyme-linked immunosorbent assay of sulfamethoxypyridazine. J Agric Food Chem ,58: 3284-3289. |
| [13] | Husain Q (2009) Peroxidase mediated decolorization and remediation of wastewater containing industrial dyes: a review. Rev Environ Sci Bio/Technology ,9: 117-140. |
| [14] | Mceldoon JP, Dordick JS (1996) Unusual Thermal Stability of Soybean Peroxidase. Biotechnol Prog ,7938: 555-558. |
| [15] | Henriksen A, Mirza O, Indiani C, et al. (2001) Structure of soybean seed coat peroxidase: A plant peroxidase with unusual stability and haem-apoprotein interactions. Protein Sci ,10: 108-115. |
| [16] | Kamal JKA, Behere DV (2002) Thermal and Conformational Stability of Seed Coat Soybean Peroxidase. Biochemistry ,41: 9034-9042. |
| [17] | Gillikin JW, Graham JS (1991) Purification and Developmental Analysis of the Major Anionic Peroxidase from the Seed Coat of Glycine max. Plant Physiol ,96: 214-220. |
| [18] | Hailu G, Weersink A, Cahlík F (2010) Examining the Prospects for Commercialization of Soybean Peroxidase. AgBioForum ,13: 263-273. |
| [19] | Franco FL, Batista-Viera F, Carlsson J (2004) Preparation of high-density Concanavalin A adsorbent and its use for rapid, high-yield purification of peroxidase from horseradish roots. J Chromatogr B Analyt Technol Biomed Life Sci ,803: 237-241. |
| [20] | Jungbauer A (1993) Review Preparative chromatography of biomolecules. J Chromatogr ,639: 3-16. |
| [21] | Patchornik G, Albeck A (2005) Free nonimmobilized ligands as a tool for purification of proteins. Bioconjug Chem ,16: 1310-1315. |
| [22] | Lam SK, Ng TB (2011) Lectins: production and practical applications. Appl Microbiol Biotechnol ,89: 45-55. |
| [23] | Lee LY, Hincapie M, Packer N, et al. (2012) An optimized approach for enrichment of glycoproteins from cell culture lysates using native multi-lectin affinity chromatography. J Sep Sci ,35: 2445-2452. |
| [24] | Hilbrig F, Stocker G, Schläppi J-M, et al. (2006) Utilization of Group Specific Ligands in the Downstream Processing of Proteins by Affinity Precipitation. Food Bioprod Process ,84: 28-36. |
| [25] | Gottschalk U (2005) Downstream Processing of Monoclonal Antibodies: from High Dilution to High Purity. BioPharm Int ,18: 42-58. |
| [26] | Linné-Larsson E, Galaev I, Lindahl L, et al. (1996) Affinity precipitation of Concanavalin A with p-aminophenyl- α-D-glucopyranoside modified Eudragit S-100. Bioseparation ,6: 273-282. |
| [27] | Urry DW, Gowda DC, Parker TM, et al. (1992) Hydrophobicity scale for proteins based on inverse temperature transitions. Biopolymers ,32: 1243-1250. |
| [28] | Fong B, Wu W, Wood DW (2009) Optimization of ELP-intein mediated protein purification by salt substitution. Protein Expr Purif ,66: 198-202. |
| [29] | Madan B, Chaudhary G, Cramer SM, et al. (2013) ELP-z and ELP-zz capturing scaffolds for the purification of immunoglobulins by affinity precipitation. J Biotechnol ,163: 10-16. |
| [30] | Arnold L, Chen R (2014) Novel thermo-responsive fucose binding ligands for glycoprotein purification by affinity precipitation. Biotechnol Bioeng ,111: 413-417. |
| [31] | Liu J, Zhang Y, Qiu L, et al. (2005) A Novel Process of Purifying Soybean Hull Peroxidase. Chem Biochem Eng Q ,19: 199-205. |
| [32] | Gray JS, Yang BY, Hull SR, et al. (1996) The glycans of soybean peroxidase. Glycobiology. ,6: 23-32. |
| [33] | Sessa DJ (2004) Processing of soybean hulls to enhance the distribution and extraction of value-added proteins. J Sci Food Agric ,84: 75-82. |
| [34] | Meyer DE, Chilkoti A (1999) Purification of recombinant proteins by fusion with thermally-responsive polypeptides. Nat Biotechnol ,17: 1112-1115. |
| [35] | Meyer DE, Trabbic-Carlson K, Chilkoti A (2001) Protein purification by fusion with an environmentally responsive elastin-like polypeptide: effect of polypeptide length on the purification of thioredoxin. Biotechnol Prog ,17: 720-728. |
| [36] | Banki MR, Feng LA, Wood DW (2005) Simple bioseparations using self-cleaving elastin-like polypeptide tags. Nat Methods ,2: 659-661. |
| [37] | Wood DW (2010) Non-Chromatographic Recombinant Protein Purification by Self-Cleaving Purification Tags. Sep Sci Technol ,45: 2345-2357. |
| [38] | Ge X, Yang DSC, Trabbic-carlson K, et al. (2005) Self-Cleavable Stimulus Responsive Tags for Protein Purification without Chromatography. J Am Chem Soc ,127: 11228-11229. |
| [39] | Tian L, Sun SSM (2011) A cost-effective ELP-intein coupling system for recombinant protein purification from plant production platform. PLoS One ,6: 24183. |
| [40] | Shimazu M, Mulchandani A, Chen W (2003) Thermally triggered purification and immobilization of elastin-OPH fusions. Biotechnol Bioeng ,81: 74-79. |
| [41] | Liu F, Tsai S-L, Madan B, et al. (2012) Engineering a high-affinity scaffold for non-chromatographic protein purification via intein-mediated cleavage. Biotechnol Bioeng ,109: 2829-2835. |
| [42] | Kostlanova N, Mitchell EP, Lortat-Jacob H, et al. (2005) The fucose-binding lectin from Ralstonia solanacearum - A new type of beta-propeller architecture formed by oligomerization and interacting with fucoside, fucosyllactose, and plant xyloglucan. J Biol Chem ,280: 27839-27849. |
| [43] | Mammen M, Choi S-K, Whitesides GM (1998) Polyvalent Interactions in Biological Systems: Implications for Design and Use of Multivalent Ligands and Inhibitors. Angew Chemie Int Ed ,37: 2754-2794. |
| [44] | Wang L-X, Amin MN (2014) Chemical and chemoenzymatic synthesis of glycoproteins for deciphering functions. Chem Biol ,21: 51-66. |
| [45] | Sheth RD, Jin M, Bhut BV, et al. (2014) Affinity precipitation of a monoclonal antibody from an industrial harvest feedstock using an ELP-Z stimuli responsive biopolymer. Biotechnol Bioeng ,111: 1595-1603. |
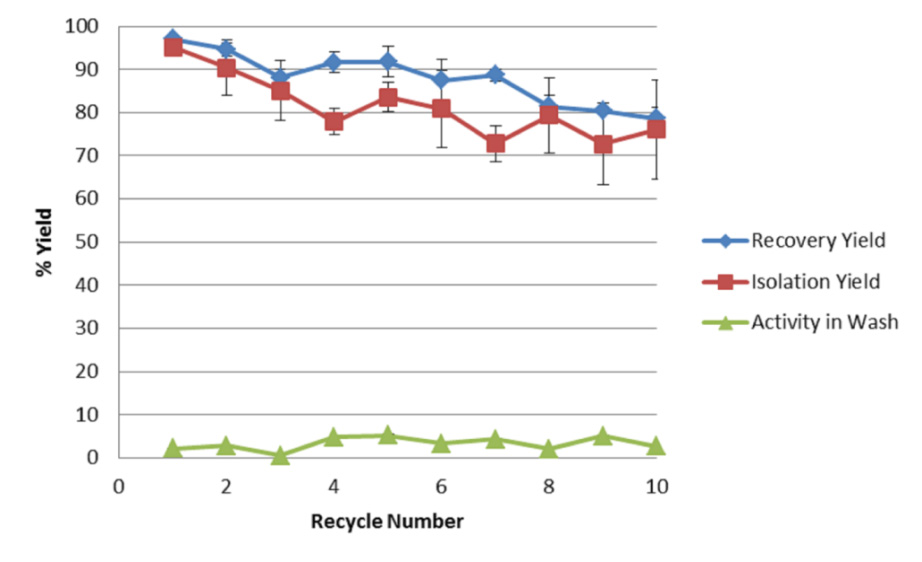
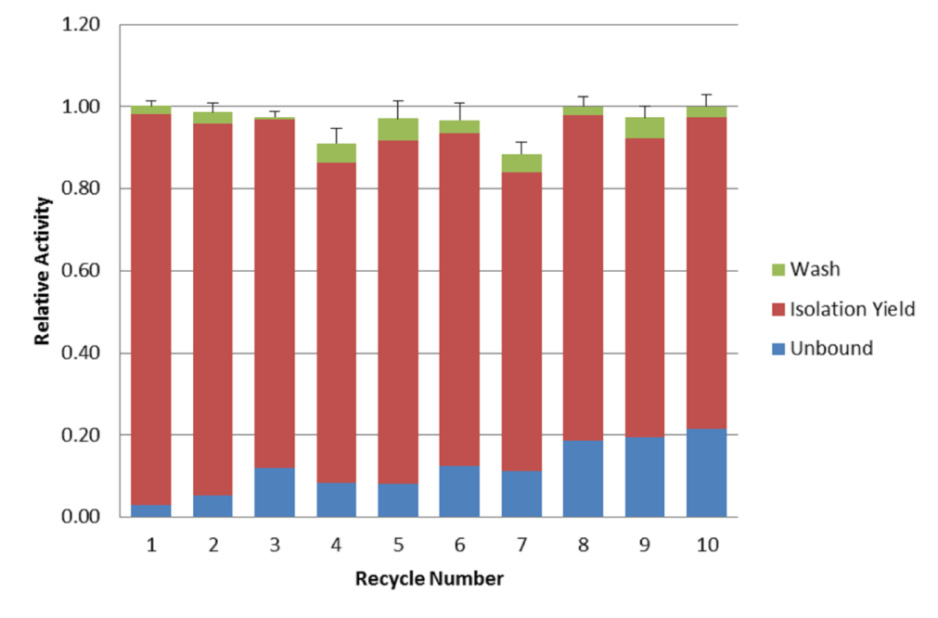
Figures(6) / Tables(2)

Lindsay Arnold, Rachel Chen. One-step non-chromatography purification of a low abundant fucosylated protein from complex plant crude extract[J]. AIMS Bioengineering, 2015, 2(3): 249-263. doi: 10.3934/bioeng.2015.3.249







 DownLoad:
DownLoad: