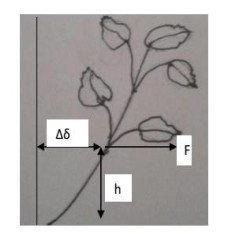
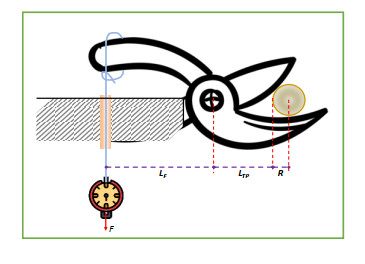
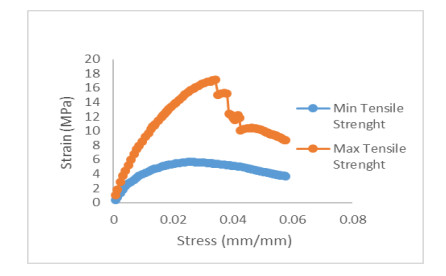
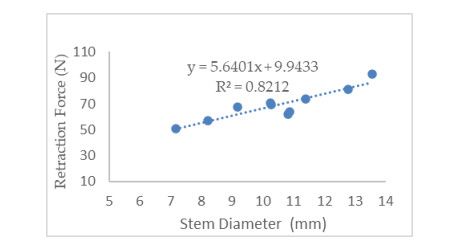
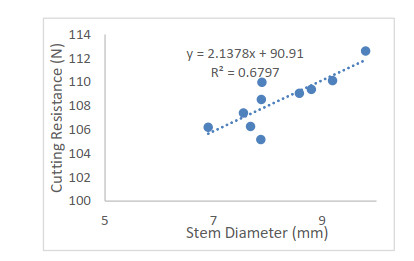
Patchouli plant characteristics are the basis that must be ascertain to design a patchouli harvester machine. The objective of this study was to determine the physical and mechanical characteristics of patchouli plants that can be used in the design of patchouli harvester machines. The data were collected by doing observation and measurement in the field and laboratory. Patchouli plants used with water content from 30 to 40% at the age of harvest 8 months after planting. Various physical characteristics of patchouli indicated the average value of plant height 112.7 cm, clump circumference 1.67 m, main stem diameter 9.88 mm, the clump diameter at a height of 20 cm from the surface of the land 16.5 cm, mass/clump 1.48 kg, and bulk density 0.11 kg/L. Physical characteristics play an important role in choosing equipment and the main dimension in designing a harvester machine. Determination of mechanical characteristics showed that the tensile strength value of patchouli was from 5.7 to 17.15 MPa with a modulus of elasticity from 0.12 to 0.5 GPa. The forces required for the design of the harvester machine such as the laying force, the retraction force and patchouli's cutting resistance achieved an average value of 6.19 N, 68.89 N, 108.47 N, respectively. The mean angle of repose was 48° at 32 % moisture content and friction coefficient was 0.71. The results obtained about the physical and mechanical characteristics of patchouli plants can be useful in providing information that helps in designing patchouli harvesting machines.
Citation: Andriani Lubis, Tineke Mandang, Wawan Hermawan, Sutrisno. Study of the physical and mechanical characteristics of patchouli plants[J]. AIMS Agriculture and Food, 2021, 6(2): 525-537. doi: 10.3934/agrfood.2021030
Patchouli plant characteristics are the basis that must be ascertain to design a patchouli harvester machine. The objective of this study was to determine the physical and mechanical characteristics of patchouli plants that can be used in the design of patchouli harvester machines. The data were collected by doing observation and measurement in the field and laboratory. Patchouli plants used with water content from 30 to 40% at the age of harvest 8 months after planting. Various physical characteristics of patchouli indicated the average value of plant height 112.7 cm, clump circumference 1.67 m, main stem diameter 9.88 mm, the clump diameter at a height of 20 cm from the surface of the land 16.5 cm, mass/clump 1.48 kg, and bulk density 0.11 kg/L. Physical characteristics play an important role in choosing equipment and the main dimension in designing a harvester machine. Determination of mechanical characteristics showed that the tensile strength value of patchouli was from 5.7 to 17.15 MPa with a modulus of elasticity from 0.12 to 0.5 GPa. The forces required for the design of the harvester machine such as the laying force, the retraction force and patchouli's cutting resistance achieved an average value of 6.19 N, 68.89 N, 108.47 N, respectively. The mean angle of repose was 48° at 32 % moisture content and friction coefficient was 0.71. The results obtained about the physical and mechanical characteristics of patchouli plants can be useful in providing information that helps in designing patchouli harvesting machines.

| [1] | Juniardi MTH, dan Hadayani MA (2015) Analisis produksi nilam dan nilai tambah penyulingan minyak atsiri di kecamatan banawa selatan kabupaten donggala. JSTT 4: 68-78. |
| [2] |
Swamy MK, Sinniah UR (2015) A comprehensive review on the phytochemical constituents and pharmacological activities of Pogostemon cablin Benth: An aromatic medicinal plant of industrial importance. Molecules 20: 8521-8547. doi: 10.3390/molecules20058521

|
| [3] |
Lubbe A, Verpoorte R (2011) Cultivation of medicinal and aromatic plants for specialty industrial materials. Ind Crops Prod 34: 785-801. doi: 10.1016/j.indcrop.2011.01.019
|
| [4] | Lisma Y (2018) Strategi pengembangan agroindustri nilam (studi kasus: koperasi industri nilam aceh di kabupaten aceh barat). Theses. IPB University. |
| [5] | Supriono, Susanti TA (2016) Kualitas minyak atsiri nilam dari metode pengecilan ukuran pada penyulingan Tanaman Nilam (Pogostemom cablin Benth). Prosiding seminar kimia. Available form: http://jurnal.kimia.fmipa.unmul.ac.id/index.php/prosiding/article/view/151. |
| [6] |
Li ZG, Li PP, Liu JZ (2011) Physical and mechanical properties of tomato fruits as related to robot's harvesting. J Food Eng 103: 170-178. doi: 10.1016/j.jfoodeng.2010.10.013
|
| [7] | Soyoye BO, Ademosun OC, Agbetoye LAS (2018) Determination of some physical and mechanical properties of soybean and maize in relation to planter design. Agric Eng Int CIGR J 20: 81-89. |
| [8] | Cao Z, Jin X, Liao QX (2011) Experimental research on physical and mechanical parameters of matured bottom stalk of the reed. Int J Agric Biol Eng 4: 36-42. |
| [9] | Haryudin W (2014) Morphological characteristics, production and quality of 15 patchouli accessions. Bul. Littro 25: 1-10. |
| [10] | Singh HJ, De D, Sahoo PK (2014) Physical properties of soybean cultivated in NEH region of India. Agric Eng Int CIGR J 16: 55-59. |
| [11] |
Chattopadhyay PS, Pandey KP (1999) Mechanical properties of sorghum stalk in relation to quasi-static deformation. J Agric Eng Res 73: 199-206. doi: 10.1006/jaer.1999.0406
|
| [12] | Milani E, Razavi MSA, Koocheki A, et al. (2007) Moisture dependent physical properties of cucurbit seeds. Int Agrophys 21: 157-168. |
| [13] |
Kumar GVP, Raheman H (2011) Development of a walk-behind type hand tractor powered vegetable transplanter for paper pot seedlings. Biosyst Eng 110: 189-197. doi: 10.1016/j.biosystemseng.2011.08.001
|
| [14] | Horia MAE, El-Sahhar EA, Mostafa MM, et al. (2008) A developed machine to harvest carrot crop. Misr J Ag Eng 25: 1163-1173. |
| [15] |
Li ZJ, Lv K, Wang YQ, et al. (2015) Multi-scale engineering properties of tomato fruits related to harvesting, simulation and textural evaluation. LWT-Food Sci Technol 61: 444-451. doi: 10.1016/j.lwt.2014.12.018
|
| [16] |
Ince A, Urluay SU, Guzel E, et al. (2005) Bending and shearing characteristics of sunflower stalk residue. Biosyst Eng 92: 175-181. doi: 10.1016/j.biosystemseng.2005.07.003
|
| [17] | Handayani S (2007) Pengujian sifat mekakanik bambu (metode pengawetan dengan boraks). Jurnal Teknik Sipil and Perencanaan 9: 43-54. |
| [18] | Intara YI, dan Banun DP (2012) Studi sifat fisik dan mekanik parenkhim pelepah daun kelapa sawit untuk pemanfaatan sebagai bahan anyaman. Agrointek 6: 36-44. |
| [19] | Bahtiar ET, Nugroho N, Surjokusumo S, et al. (2016) Pengaruh komponen kimia dan ikatan pembuluh terhadap kekuatan tarik bambu. Jurnal Teknik Sipil 23: 31-40. |
| [20] |
Özbek O, Seflek AY, Carman K (2009) Some mechanical properties of safflower stalk. Appl Eng Agric 25: 619-626. doi: 10.13031/2013.27452
|
| [21] |
Jahanbakhshi A, Abbaspour-Gilandeh Y, Gundoshmian TM (2018) Determination of physical and mechanical properties of carrot in order to reduce waste during harvesting and post-harvesting. Food Sci Nutr 6: 1898-1903. doi: 10.1002/fsn3.760
|
| [22] |
Galedar MN, Jafaria A, Mohtasebia SS, et al. (2008) Effects of moisture content and level in the crop on the engineering properties of alfalfa stems. Biosyst Eng 101: 199-208. doi: 10.1016/j.biosystemseng.2008.07.006
|
| [23] | Lisyanto (2007) Evaluasi parameter desain piring pengolah tanah diputar untuk pengepras tebu lahan kering. Theses. IPB University. |
| [24] |
Igathinathane C, Womac AR, Sokhansanj S, et al. (2009) Size reduction of high- and low-moisture corn stalks by linear knife grid system. Biomass Bioenerg 33: 547-557. doi: 10.1016/j.biombioe.2008.09.004
|
| [25] | Dange AR, Thakare SK, Rao IB (2011) Cutting energy and force as required for Pigeon pea stems. J Agric Technol 7: 1485-1493. |
| [26] |
Igathinathane C, Womac AR, Sokhansanj S (2010) Corn stalk orientation effect on mechanical cutting. Biosyst Eng 107: 97-106. doi: 10.1016/j.biosystemseng.2010.07.005
|
| [27] |
Johnson PC, Clementson CL, Mathanker SK, et al. (2012) Cutting energy characteristics of Miscanthus x giganteus stems with varying oblique angle and cutting speed. Biosyst Eng 112: 42-48. doi: 10.1016/j.biosystemseng.2012.02.003
|
| [28] | Dauda SM, Ahmad DA. Khalina A, et al. (2014) Physical and mechanical properties of kenaf stems at varying moisture contents. Agric Agric Sci Procedia 2: 370-374. |
| [29] | Seifi MR, Alimardani R (2010) The moisture content effect on some physical and mechanical properties of corn (Sc 704). J Agric Sci 2: 125-134. |
| [30] | El-Fawal YA, Tawfik MA, El-Shal AM (2009) Study on physical and engineering properties for grains of some field crops. Misr J Ag Eng 26: 1933-1951. |
| [31] | Davies R, Yusuf DD (2017) Studies of physical and mechanical properties of velvet tamarind. MAYFEB J Agric Sci 2: 36-43. |

Figures(6) / Tables(4)

Andriani Lubis, Tineke Mandang, Wawan Hermawan, Sutrisno. Study of the physical and mechanical characteristics of patchouli plants[J]. AIMS Agriculture and Food, 2021, 6(2): 525-537. doi: 10.3934/agrfood.2021030







 DownLoad:
DownLoad: