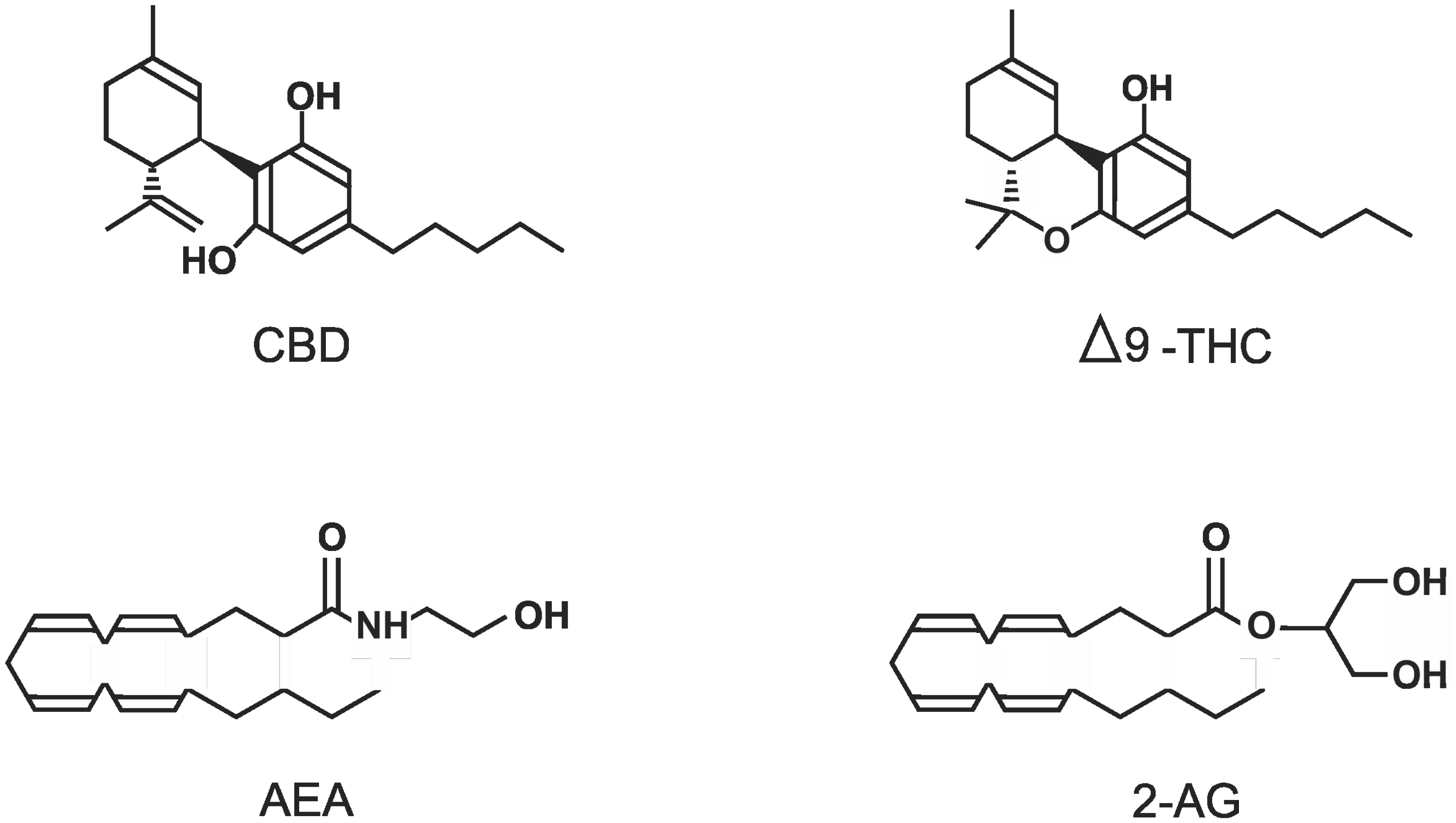
Marijuana has been used by humans for thousands of years for both medicinal and recreational purposes. This included the treatment of pain, inflammation, seizures, and nausea. In the 1960s, the structure of the principal psychoactive ingredient Δ9-tetrahydrocannabinol was determined, and over the next few decades, two cannabinoid receptors were characterized along with the human endocannabinoid system and what it affects. This includes metabolism, the cardiovascular and reproductive systems, and it is involved in such conditions as inflammation, cancer, glaucoma, and liver and musculoskeletal disorders. In the central nervous system, the endocannabinoid system has been linked to appetite, learning, memory, and conditions such as depression, anxiety, schizophrenia, stroke, multiple sclerosis, neurodegeneration, addiction, and epilepsy. It was the profound effectiveness of cannabidiol, a non-psychoactive ingredient of marijuana, to relieve the symptoms of Dravet syndrome, a severe form of childhood epilepsy, that recently helped spur marijuana research. This has helped substantially to change society's attitude towards this potential source of useful drugs. However, research has also revealed that the actions of endocannabinoids, such as anandamide and 2-arachidonoylglycerol, and the phytocannabinoids, tetrahydrocannabinol and cannabidiol, were not just due to interactions with the two cannabinoid receptors but by acting directly on many other targets including various G-protein receptors and cation channels, such as the transient receptor potential channels for example. This mini-review attempts to survey the effects of these 4 important cannabinoids on these currently identified targets.
Citation: Nicholas J. D. Wright. A review of the direct targets of the cannabinoids cannabidiol, Δ9-tetrahydrocannabinol, N-arachidonoylethanolamine and 2-arachidonoylglycerol[J]. AIMS Neuroscience, 2024, 11(2): 144-165. doi: 10.3934/Neuroscience.2024009
Marijuana has been used by humans for thousands of years for both medicinal and recreational purposes. This included the treatment of pain, inflammation, seizures, and nausea. In the 1960s, the structure of the principal psychoactive ingredient Δ9-tetrahydrocannabinol was determined, and over the next few decades, two cannabinoid receptors were characterized along with the human endocannabinoid system and what it affects. This includes metabolism, the cardiovascular and reproductive systems, and it is involved in such conditions as inflammation, cancer, glaucoma, and liver and musculoskeletal disorders. In the central nervous system, the endocannabinoid system has been linked to appetite, learning, memory, and conditions such as depression, anxiety, schizophrenia, stroke, multiple sclerosis, neurodegeneration, addiction, and epilepsy. It was the profound effectiveness of cannabidiol, a non-psychoactive ingredient of marijuana, to relieve the symptoms of Dravet syndrome, a severe form of childhood epilepsy, that recently helped spur marijuana research. This has helped substantially to change society's attitude towards this potential source of useful drugs. However, research has also revealed that the actions of endocannabinoids, such as anandamide and 2-arachidonoylglycerol, and the phytocannabinoids, tetrahydrocannabinol and cannabidiol, were not just due to interactions with the two cannabinoid receptors but by acting directly on many other targets including various G-protein receptors and cation channels, such as the transient receptor potential channels for example. This mini-review attempts to survey the effects of these 4 important cannabinoids on these currently identified targets.

| [1] |
Iversen L (2003) Cannabis and the brain. Brain 126: 1252-1270. https://doi.org/10.1093/brain/awg143 
|
| [2] | Mechoulam R (1986) The Pharmacohistory of Cannabis Sativa. Cannabinoids As Therapeutic Agents . Chapman and Hall/CRC pp. 1-20. https://doi.org/10.1201/9780429260667-1 |
| [3] |
Gaoni Y, Mechoulam R (1964) Isolation, Structure, and Partial Synthesis of an Active Constituent of Hashish. J Am Chem Soc 86: 1646-1647. https://doi.org/10.1021/ja01062a046
|
| [4] | Devane WA, Dysarz FA, Johnson MR, et al. (1988) Determination and characterization of a cannabinoid receptor in rat brain. Mol Pharmacol 34: 605-613. |
| [5] |
Matsuda LA, Lolait SJ, Brownstein MJ, et al. (1990) Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 346: 561-564. https://doi.org/10.1038/346561a0
|
| [6] |
Munro S, Thomas KL, Abu-Shaar M (1993) Molecular characterization of a peripheral receptor for cannabinoids. Nature 365: 61-65. https://doi.org/10.1038/365061a0
|
| [7] |
Pertwee RG, Howlett AC, Abood ME, et al. (2010) International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: beyond CB1 and CB2. Pharmacol Rev 62: 588-631. https://doi.org/10.1124/pr.110.003004
|
| [8] |
Devane WA, Hanuš L, Breuer A, et al. (1992) Isolation and Structure of a Brain Constituent That Binds to the Cannabinoid Receptor. Science 258: 1946-1949. https://doi.org/10.1126/science.1470919
|
| [9] |
Mechoulam R, Ben-Shabat S, Hanus L, et al. (1995) Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol 50: 83-90. https://doi.org/10.1016/0006-2952(95)00109-D
|
| [10] |
Pacher P, Bátkai S, Kunos G (2006) The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev 58: 389-462. https://doi.org/10.1124/pr.58.3.2
|
| [11] |
Sugiura T, Kondo S, Sukagawa A, et al. (1995) 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys Res Commun 215: 89-97. https://doi.org/10.1006/bbrc.1995.2437
|
| [12] |
Kano M, Ohno-Shosaku T, Hashimotodani Y, et al. (2009) Endocannabinoid-mediated control of synaptic transmission. Physiol Rev 89: 309-380. https://doi.org/10.1152/physrev.00019.2008
|
| [13] |
Di Marzo V, Stella N, Zimmer A (2015) Endocannabinoid signalling and the deteriorating brain. Nat Rev Neurosci 16: 30-42. https://doi.org/10.1038/nrn3876
|
| [14] |
Maccarrone M, Bab I, Bíró T, et al. (2015) Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol Sci 36: 277-296. https://doi.org/10.1016/j.tips.2015.02.008
|
| [15] |
Miller LK, Devi LA (2011) The highs and lows of cannabinoid receptor expression in disease: mechanisms and their therapeutic implications. Pharmacol Rev 63: 461-470. https://doi.org/10.1124/pr.110.003491
|
| [16] |
Adams R, Hunt M, Clark JH (1940) Structure of Cannabidiol, a Product Isolated from the Marihuana Extract of Minnesota Wild Hemp. I. J Am Chem Soc 62: 196-200. https://doi.org/10.1021/ja01858a058
|
| [17] |
Jacob A, Todd AR (1940) Cannabidiol and Cannabol, Constituents of Cannabis indica Resin. Nature 145: 350-350. https://doi.org/10.1038/145350a0
|
| [18] |
Mechoulam R, Shvo Y (1963) Hashish. I. The structure of cannabidiol. Tetrahedron 19: 2073-2078. https://doi.org/10.1016/0040-4020(63)85022-X
|
| [19] |
Kimura M, Okamoto K (1970) Distribution of tetrahydrocannabinolic acid in fresh wild cannabis. Experientia 26: 819-820. https://doi.org/10.1007/BF02114192
|
| [20] |
Taura F, Sirikantaramas S, Shoyama Y, et al. (2007) Phytocannabinoids in Cannabis sativa: recent studies on biosynthetic enzymes. Chem Biodivers 4: 1649-1663. https://doi.org/10.1002/cbdv.200790145
|
| [21] |
Araque A, Castillo PE, Manzoni OJ, et al. (2017) Synaptic functions of endocannabinoid signaling in health and disease. Neuropharmacology 124: 13-24. https://doi.org/10.1016/j.neuropharm.2017.06.017
|
| [22] |
Hua T, Vemuri K, Nikas SP, et al. (2017) Crystal structures of agonist-bound human cannabinoid receptor CB(1). Nature 547: 468-471. https://doi.org/10.1038/nature23272
|
| [23] |
Hua T, Vemuri K, Pu M, et al. (2016) Crystal Structure of the Human Cannabinoid Receptor CB1. Cell 167: 750-762.e714. https://doi.org/10.1016/j.cell.2016.10.004
|
| [24] |
Shao Z, Yin J, Chapman K, et al. (2016) High-resolution crystal structure of the human CB1 cannabinoid receptor. Nature 540: 602-606. https://doi.org/10.1038/nature20613
|
| [25] |
González-Mariscal I, Krzysik-Walker SM, Doyle ME, et al. (2016) Human CB1 Receptor Isoforms, present in Hepatocytes and β-cells, are Involved in Regulating Metabolism. Sci Rep 6: 33302. https://doi.org/10.1038/srep33302
|
| [26] |
Clapper JR, Moreno-Sanz G, Russo R, et al. (2010) Anandamide suppresses pain initiation through a peripheral endocannabinoid mechanism. Nat Neurosci 13: 1265-1270. https://doi.org/10.1038/nn.2632
|
| [27] |
Price TJ, Helesic G, Parghi D, et al. (2003) The neuronal distribution of cannabinoid receptor type 1 in the trigeminal ganglion of the rat. Neuroscience 120: 155-162. https://doi.org/10.1016/S0306-4522(03)00333-6
|
| [28] |
Tam J, Trembovler V, Di Marzo V, et al. (2008) The cannabinoid CB1 receptor regulates bone formation by modulating adrenergic signaling. Faseb J 22: 285-294. https://doi.org/10.1096/fj.06-7957com
|
| [29] |
Veress G, Meszar Z, Muszil D, et al. (2013) Characterisation of cannabinoid 1 receptor expression in the perikarya, and peripheral and spinal processes of primary sensory neurons. Brain Struct Funct 218: 733-750. https://doi.org/10.1007/s00429-012-0425-2
|
| [30] |
Howlett AC, Barth F, Bonner TI, et al. (2002) International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol Rev 54: 161-202. https://doi.org/10.1124/pr.54.2.161
|
| [31] |
Demuth DG, Molleman A (2006) Cannabinoid signalling. Life Sci 78: 549-563. https://doi.org/10.1016/j.lfs.2005.05.055
|
| [32] |
Rhee M-H, Bayewitch M, Avidor-Reiss T, et al. (1998) Cannabinoid Receptor Activation Differentially Regulates the Various Adenylyl Cyclase Isozymes. J Neurochem 71: 1525-1534. https://doi.org/10.1046/j.1471-4159.1998.71041525.x
|
| [33] |
Navarrete M, Araque A (2008) Endocannabinoids mediate neuron-astrocyte communication. Neuron 57: 883-893. https://doi.org/10.1016/j.neuron.2008.01.029
|
| [34] |
Brown SP, Safo PK, Regehr WG (2004) Endocannabinoids inhibit transmission at granule cell to Purkinje cell synapses by modulating three types of presynaptic calcium channels. J Neurosci 24: 5623-5631. https://doi.org/10.1523/JNEUROSCI.0918-04.2004
|
| [35] |
Szabó GG, Lenkey N, Holderith N, et al. (2014) Presynaptic calcium channel inhibition underlies CB1 cannabinoid receptor-mediated suppression of GABA release. J Neurosci 34: 7958-7963. https://doi.org/10.1523/JNEUROSCI.0247-14.2014
|
| [36] |
Guo J, Ikeda SR (2004) Endocannabinoids modulate N-type calcium channels and G-protein-coupled inwardly rectifying potassium channels via CB1 cannabinoid receptors heterologously expressed in mammalian neurons. Mol Pharmacol 65: 665-674. https://doi.org/10.1124/mol.65.3.665
|
| [37] |
Mackie K, Lai Y, Westenbroek R, et al. (1995) Cannabinoids activate an inwardly rectifying potassium conductance and inhibit Q-type calcium currents in AtT20 cells transfected with rat brain cannabinoid receptor. J Neurosci 15: 6552-6561. https://doi.org/10.1523/JNEUROSCI.15-10-06552.1995
|
| [38] |
Galve-Roperh I, Rueda D, Gómez del Pulgar T, et al. (2002) Mechanism of extracellular signal-regulated kinase activation by the CB(1) cannabinoid receptor. Mol Pharmacol 62: 1385-1392. https://doi.org/10.1124/mol.62.6.1385
|
| [39] |
Gómez del Pulgar T, Velasco G, Guzmán M (2000) The CB1 cannabinoid receptor is coupled to the activation of protein kinase B/Akt. Biochem J 347: 369-373. https://doi.org/10.1042/bj3470369
|
| [40] |
Liu QR, Pan CH, Hishimoto A, et al. (2009) Species differences in cannabinoid receptor 2 (CNR2 gene): identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav 8: 519-530. https://doi.org/10.1111/j.1601-183X.2009.00498.x
|
| [41] |
Bouaboula M, Poinot-Chazel C, Bourrié B, et al. (1995) Activation of mitogen-activated protein kinases by stimulation of the central cannabinoid receptor CB1. Biochem J 312: 637-641. https://doi.org/10.1042/bj3120637
|
| [42] |
Bouaboula M, Poinot-Chazel C, Marchand J, et al. (1996) Signaling pathway associated with stimulation of CB2 peripheral cannabinoid receptor. Involvement of both mitogen-activated protein kinase and induction of Krox-24 expression. Eur J Biochem 237: 704-711. https://doi.org/10.1111/j.1432-1033.1996.0704p.x
|
| [43] |
Viscomi MT, Oddi S, Latini L, et al. (2009) Selective CB2 receptor agonism protects central neurons from remote axotomy-induced apoptosis through the PI3K/Akt pathway. J Neurosci 29: 4564-4570. https://doi.org/10.1523/JNEUROSCI.0786-09.2009
|
| [44] |
McPartland JM, Glass M, Pertwee RG (2007) Meta-analysis of cannabinoid ligand binding affinity and receptor distribution: interspecies differences. Br J Pharmacol 152: 583-593. https://doi.org/10.1038/sj.bjp.0707399
|
| [45] |
Pertwee RG, Ross RA, Craib SJ, et al. (2002) (-)-Cannabidiol antagonizes cannabinoid receptor agonists and noradrenaline in the mouse vas deferens. Eur J Pharmacol 456: 99-106. https://doi.org/10.1016/S0014-2999(02)02624-9
|
| [46] |
Thomas A, Baillie GL, Phillips AM, et al. (2007) Cannabidiol displays unexpectedly high potency as an antagonist of CB1 and CB2 receptor agonists in vitro. Br J Pharmacol 150: 613-623. https://doi.org/10.1038/sj.bjp.0707133
|
| [47] |
Laprairie RB, Bagher AM, Kelly ME, et al. (2015) Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br J Pharmacol 172: 4790-4805. https://doi.org/10.1111/bph.13250
|
| [48] |
Morales P, Goya P, Jagerovic N, et al. (2016) Allosteric Modulators of the CB(1) Cannabinoid Receptor: A Structural Update Review. Cannabis Cannabinoid Res 1: 22-30. https://doi.org/10.1089/can.2015.0005
|
| [49] |
Tham M, Yilmaz O, Alaverdashvili M, et al. (2019) Allosteric and orthosteric pharmacology of cannabidiol and cannabidiol-dimethylheptyl at the type 1 and type 2 cannabinoid receptors. Br J Pharmacol 176: 1455-1469. https://doi.org/10.1111/bph.14440
|
| [50] |
Pertwee RG (1999) Pharmacology of cannabinoid receptor ligands. Curr Med Chem 6: 635-664. https://doi.org/10.2174/0929867306666220401124036
|
| [51] | Pertwee RG (2005) Pharmacological actions of cannabinoids. Handb Exp Pharmacol : 1-51. https://doi.org/10.1007/3-540-26573-2_1 |
| [52] |
Di Marzo V, De Petrocellis L (2012) Why do cannabinoid receptors have more than one endogenous ligand?. Philos Trans R Soc Lond B Biol Sci 367: 3216-3228. https://doi.org/10.1098/rstb.2011.0382
|
| [53] |
Brown KJ, Laun AS, Song ZH (2017) Cannabidiol, a novel inverse agonist for GPR12. Biochem Biophys Res Commun 493: 451-454. https://doi.org/10.1016/j.bbrc.2017.09.001
|
| [54] |
Laun AS, Shrader SH, Brown KJ, et al. (2019) GPR3, GPR6, and GPR12 as novel molecular targets: their biological functions and interaction with cannabidiol. Acta Pharmacol Sin 40: 300-308. https://doi.org/10.1038/s41401-018-0031-9
|
| [55] |
Laun AS, Shrader SH, Song Z-H (2018) Novel inverse agonists for the orphan G protein-coupled receptor 6. Heliyon 4: e00933. https://doi.org/10.1016/j.heliyon.2018.e00933
|
| [56] |
Gantz I, Muraoka A, Yang Y-K, et al. (1997) Cloning and Chromosomal Localization of a Gene (GPR18) Encoding a Novel Seven Transmembrane Receptor Highly Expressed in Spleen and Testis. Genomics 42: 462-466. https://doi.org/10.1006/geno.1997.4752
|
| [57] |
Penumarti A, Abdel-Rahman AA (2014) The Novel Endocannabinoid Receptor GPR18 Is Expressed in the Rostral Ventrolateral Medulla and Exerts Tonic Restraining Influence on Blood Pressure. J Pharmacol Exp Ther 349: 29-38. https://doi.org/10.1124/jpet.113.209213
|
| [58] |
McHugh D, Roskowski D, Xie S, et al. (2014) Δ(9)-THC and N-arachidonoyl glycine regulate BV-2 microglial morphology and cytokine release plasticity: implications for signaling at GPR18. Front Pharmacol 4: 162. https://doi.org/10.3389/fphar.2013.00162
|
| [59] |
McHugh D, Page J, Dunn E, et al. (2012) Δ(9)-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br J Pharmacol 165: 2414-2424. https://doi.org/10.1111/j.1476-5381.2011.01497.x
|
| [60] |
Obara Y, Ueno S, Yanagihata Y, et al. (2011) Lysophosphatidylinositol Causes Neurite Retraction via GPR55, G13 and RhoA in PC12 Cells. PLOS ONE 6: e24284. https://doi.org/10.1371/journal.pone.0024284
|
| [61] |
Ryberg E, Larsson N, Sjögren S, et al. (2007) The orphan receptor GPR55 is a novel cannabinoid receptor. Brit J Pharmacol 152: 1092-1101. https://doi.org/10.1038/sj.bjp.0707460
|
| [62] |
Lauckner JE, Jensen JB, Chen H-Y, et al. (2008) GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc Natl Acad Sci 105: 2699-2704. https://doi.org/10.1073/pnas.0711278105
|
| [63] |
Whyte LS, Ryberg E, Sims NA, et al. (2009) The putative cannabinoid receptor GPR55 affects osteoclast function in vitro and bone mass in vivo. Proc Natl Acad Sci U S A 106: 16511-16516. https://doi.org/10.1073/pnas.0902743106
|
| [64] |
Sharir H, Console-Bram L, Mundy C, et al. (2012) The endocannabinoids anandamide and virodhamine modulate the activity of the candidate cannabinoid receptor GPR55. J Neuroimmune Pharmacol 7: 856-865. https://doi.org/10.1007/s11481-012-9351-6
|
| [65] | Corbett AD, Henderson G, McKnight AT, et al. (2006) 75 years of opioid research: the exciting but vain quest for the Holy Grail. Br J Pharmacol 147 Suppl 1: S153-162. https://doi.org/10.1038/sj.bjp.0706435 |
| [66] |
Hine B, Friedman E, Torrelio M, et al. (1975) Morphine-Dependent Rats: Blockade of Precipitated Abstinence by Tetrahydrocannabinol. Science 187: 443-445. https://doi.org/10.1126/science.1167428
|
| [67] |
Cox BM, Christie MJ, Devi L, et al. (2015) Challenges for opioid receptor nomenclature: IUPHAR Review 9. Brit J Pharmacol 172: 317-323. https://doi.org/10.1111/bph.12612
|
| [68] |
Tulunay FC, Ayhan IH, Portoghese PS, et al. (1981) Antagonism by chlornaltrexamine of some effects of Δ9-tetrahydrocannabinol in rats. Eur J Pharmacol 70: 219-224. https://doi.org/10.1016/0014-2999(81)90217-X
|
| [69] | Vaysse PJ, Gardner EL, Zukin RS (1987) Modulation of rat brain opioid receptors by cannabinoids. J Pharmacol Exp Ther 241: 534-539. |
| [70] |
Kathmann M, Flau K, Redmer A, et al. (2006) Cannabidiol is an allosteric modulator at mu- and delta-opioid receptors. Naunyn Schmiedebergs Arch Pharmacol 372: 354-361. https://doi.org/10.1007/s00210-006-0033-x
|
| [71] | Smith PB, Welch SP, Martin BR (1994) Interactions between delta 9-tetrahydrocannabinol and kappa opioids in mice. J Pharmacol Exp Ther 268: 1381-1387. |
| [72] |
Reis GML, Pacheco D, Perez AC, et al. (2009) Opioid receptor and NO/cGMP pathway as a mechanism of peripheral antinociceptive action of the cannabinoid receptor agonist anandamide. Life Sci 85: 351-356. https://doi.org/10.1016/j.lfs.2009.06.012
|
| [73] |
Haller VL, Stevens DL, Welch SP (2008) Modulation of opioids via protection of anandamide degradation by fatty acid amide hydrolase. Eur J Pharmacol 600: 50-58. https://doi.org/10.1016/j.ejphar.2008.08.005
|
| [74] |
Burnstock G (2017) Purinergic Signalling: Therapeutic Developments. Front Pharmacol 8. https://doi.org/10.3389/fphar.2017.00661
|
| [75] |
Cunha RA (2001) Adenosine as a neuromodulator and as a homeostatic regulator in the nervous system: different roles, different sources and different receptors. Neurochem Int 38: 107-125. https://doi.org/10.1016/S0197-0186(00)00034-6
|
| [76] |
Burnstock G (2012) Purinergic signalling: Its unpopular beginning, its acceptance and its exciting future. Bioessays 34: 218-225. https://doi.org/10.1002/bies.201100130
|
| [77] |
Carrier EJ, Auchampach JA, Hillard CJ (2006) Inhibition of an equilibrative nucleoside transporter by cannabidiol: A mechanism of cannabinoid immunosuppression. Proc Natl Acad Sci 103: 7895-7900. https://doi.org/10.1073/pnas.0511232103
|
| [78] |
Gonca E, Darıcı F (2015) The effect of cannabidiol on ischemia/reperfusion-induced ventricular arrhythmias: the role of adenosine A1 receptors. J Cardiovasc Pharmacol Ther 20: 76-83. https://doi.org/10.1177/1074248414532013
|
| [79] |
Lane JR, Beukers MW, Mulder-Krieger T, et al. (2010) The endocannabinoid 2-arachidonylglycerol is a negative allosteric modulator of the human A3 adenosine receptor. Biochem Pharmacol 79: 48-56. https://doi.org/10.1016/j.bcp.2009.07.024
|
| [80] |
Tyagi S, Gupta P, Saini AS, et al. (2011) The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J Adv Pharm Technol Res 2: 236-240. https://doi.org/10.4103/2231-4040.90879
|
| [81] |
Burstein S (2005) PPAR-γ: A nuclear receptor with affinity for cannabinoids. Life Sci 77: 1674-1684. https://doi.org/10.1016/j.lfs.2005.05.039
|
| [82] |
O'Sullivan SE (2007) Cannabinoids go nuclear: evidence for activation of peroxisome proliferator-activated receptors. Brit J Pharmacol 152: 576-582. https://doi.org/10.1038/sj.bjp.0707423
|
| [83] |
O'Sullivan SE (2016) An update on PPAR activation by cannabinoids. Br J Pharmacol 173: 1899-1910. https://doi.org/10.1111/bph.13497
|
| [84] |
Khosropoor S, Alavi MS, Etemad L, et al. (2023) Cannabidiol goes nuclear: The role of PPARγ. Phytomedicine 114: 154771. https://doi.org/10.1016/j.phymed.2023.154771
|
| [85] |
Hughes ML, Liu B, Halls ML, et al. (2015) Fatty Acid-binding Proteins 1 and 2 Differentially Modulate the Activation of Peroxisome Proliferator-activated Receptor α in a Ligand-selective Manner. J Biol Chem 290: 13895-13906. https://doi.org/10.1074/jbc.M114.605998
|
| [86] |
Alhamoruni A, Lee AC, Wright KL, et al. (2010) Pharmacological effects of cannabinoids on the Caco-2 cell culture model of intestinal permeability. J Pharmacol Exp Ther 335: 92-102. https://doi.org/10.1124/jpet.110.168237
|
| [87] |
Alhamoruni A, Wright KL, Larvin M, et al. (2012) Cannabinoids mediate opposing effects on inflammation-induced intestinal permeability. Br J Pharmacol 165: 2598-2610. https://doi.org/10.1111/j.1476-5381.2011.01589.x
|
| [88] |
Bouaboula M, Hilairet S, Marchand J, et al. (2005) Anandamide induced PPARgamma transcriptional activation and 3T3-L1 preadipocyte differentiation. Eur J Pharmacol 517: 174-181. https://doi.org/10.1016/j.ejphar.2005.05.032
|
| [89] |
Kozak KR, Gupta RA, Moody JS, et al. (2002) 15-Lipoxygenase metabolism of 2-arachidonylglycerol. Generation of a peroxisome proliferator-activated receptor alpha agonist. J Biol Chem 277: 23278-23286. https://doi.org/10.1074/jbc.M201084200
|
| [90] |
Rockwell CE, Snider NT, Thompson JT, et al. (2006) Interleukin-2 suppression by 2-arachidonyl glycerol is mediated through peroxisome proliferator-activated receptor gamma independently of cannabinoid receptors 1 and 2. Mol Pharmacol 70: 101-111. https://doi.org/10.1124/mol.105.019117
|
| [91] |
Sun Y, Alexander SP, Garle MJ, et al. (2007) Cannabinoid activation of PPAR alpha; a novel neuroprotective mechanism. Br J Pharmacol 152: 734-743. https://doi.org/10.1038/sj.bjp.0707478
|
| [92] |
Dutertre S, Becker CM, Betz H (2012) Inhibitory glycine receptors: an update. J Biol Chem 287: 40216-40223. https://doi.org/10.1074/jbc.R112.408229
|
| [93] |
Betz H, Laube B (2006) Glycine receptors: recent insights into their structural organization and functional diversity. J Neurochem 97: 1600-1610. https://doi.org/10.1111/j.1471-4159.2006.03908.x
|
| [94] |
Lynch JW (2004) Molecular structure and function of the glycine receptor chloride channel. Physiol Rev 84: 1051-1095. https://doi.org/10.1152/physrev.00042.2003
|
| [95] |
Ahrens J, Demir R, Leuwer M, et al. (2009) The Nonpsychotropic Cannabinoid Cannabidiol Modulates and Directly Activates Alpha-1 and Alpha-1-Beta Glycine Receptor Function. Pharmacology 83: 217-222. https://doi.org/10.1159/000201556
|
| [96] |
Xiong W, Cheng K, Cui T, et al. (2011) Cannabinoid potentiation of glycine receptors contributes to cannabis-induced analgesia. Nat Chem Biol 7: 296-303. https://doi.org/10.1038/nchembio.552
|
| [97] |
Hejazi N, Zhou C, Oz M, et al. (2006) Delta9-tetrahydrocannabinol and endogenous cannabinoid anandamide directly potentiate the function of glycine receptors. Mol Pharmacol 69: 991-997. https://doi.org/10.1124/mol.105.019174
|
| [98] |
Lozovaya N, Yatsenko N, Beketov A, et al. (2005) Glycine Receptors in CNS Neurons as a Target for Nonretrograde Action of Cannabinoids. J Neurosci 25: 7499-7506. https://doi.org/10.1523/JNEUROSCI.0977-05.2005
|
| [99] |
Yang Z, Aubrey KR, Alroy I, et al. (2008) Subunit-specific modulation of glycine receptors by cannabinoids and N-arachidonyl-glycine. Biochem Pharmacol 76: 1014-1023. https://doi.org/10.1016/j.bcp.2008.07.037
|
| [100] |
Alvarez LD, Alves NRC (2023) Molecular determinants of tetrahydrocannabinol binding to the glycine receptor. Proteins 91: 400-411. https://doi.org/10.1002/prot.26438
|
| [101] |
Albuquerque EX, Pereira EF, Alkondon M, et al. (2009) Mammalian nicotinic acetylcholine receptors: from structure to function. Physiol Rev 89: 73-120. https://doi.org/10.1152/physrev.00015.2008
|
| [102] |
Zoli M, Pistillo F, Gotti C (2015) Diversity of native nicotinic receptor subtypes in mammalian brain. Neuropharmacology 96: 302-311. https://doi.org/10.1016/j.neuropharm.2014.11.003
|
| [103] |
Mahgoub M, Keun-Hang SY, Sydorenko V, et al. (2013) Effects of cannabidiol on the function of α7-nicotinic acetylcholine receptors. Eur J Pharmacol 720: 310-319. https://doi.org/10.1016/j.ejphar.2013.10.011
|
| [104] |
Oz M, Al Kury L, Keun-Hang SY, et al. (2014) Cellular approaches to the interaction between cannabinoid receptor ligands and nicotinic acetylcholine receptors. Eur J Pharmacol 731: 100-–105. https://doi.org/10.1016/j.ejphar.2014.03.010
|
| [105] |
Turkanis SA, Karler R (1986) Effects of delta-9-tetrahydrocannabinol, 11 -hydroxy-delta-9-tetrahydrocannabinol and cannabidiol on neuromuscular transmission in the frog. Neuropharmacology 25: 1273-1278. https://doi.org/10.1016/0028-3908(86)90147-4
|
| [106] |
Oz M, Zhang L, Ravindran A, et al. (2004) Differential Effects of Endogenous and Synthetic Cannabinoids on α<sub>7</sub>-Nicotinic Acetylcholine Receptor-Mediated Responses in <em>Xenopus</em> Oocytes. J Pharmacol Exp Ther 310: 1152-1160. https://doi.org/10.1124/jpet.104.067751
|
| [107] |
Spivak CE, Lupica CR, Oz M (2007) The Endocannabinoid Anandamide Inhibits the Function of α4β2 Nicotinic Acetylcholine Receptors. Mol Pharmacol 72: 1024-1032. https://doi.org/10.1124/mol.107.036939
|
| [108] |
Oz M, Yang KS, Mahgoub MO (2022) Effects of cannabinoids on ligand-gated ion channels. Front Physiol 13: 1041833. https://doi.org/10.3389/fphys.2022.1041833
|
| [109] |
Hoyer D, Hannon JP, Martin GR (2002) Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol Biochem Behav 71: 533-554. https://doi.org/10.1016/S0091-3057(01)00746-8
|
| [110] |
Espejo-Porras F, Fernández-Ruiz J, Pertwee RG, et al. (2013) Motor effects of the non-psychotropic phytocannabinoid cannabidiol that are mediated by 5-HT1A receptors. Neuropharmacology 75: 155-163. https://doi.org/10.1016/j.neuropharm.2013.07.024
|
| [111] |
Russo EB, Burnett A, Hall B, et al. (2005) Agonistic properties of cannabidiol at 5-HT1a receptors. Neurochem Res 30: 1037-1043. https://doi.org/10.1007/s11064-005-6978-1
|
| [112] |
Barnes NM, Hales TG, Lummis SCR, et al. (2009) The 5-HT3 receptor – the relationship between structure and function. Neuropharmacology 56: 273-284. https://doi.org/10.1016/j.neuropharm.2008.08.003
|
| [113] |
Lummis SCR (2012) 5-HT3 Receptors*. J Biol Chem 287: 40239-40245. https://doi.org/10.1074/jbc.R112.406496
|
| [114] |
Morales M, Wang SD (2002) Differential composition of 5-hydroxytryptamine3 receptors synthesized in the rat CNS and peripheral nervous system. J Neurosci 22: 6732-6741. https://doi.org/10.1523/JNEUROSCI.22-15-06732.2002
|
| [115] |
van Hooft JA, Yakel JL (2003) 5-HT3 receptors in the CNS: 3B or not 3B?. Trends Pharmacol Sci 24: 157-160. https://doi.org/10.1016/S0165-6147(03)00051-8
|
| [116] |
Barann M, Molderings G, Brüss M, et al. (2002) Direct inhibition by cannabinoids of human 5-HT3A receptors: probable involvement of an allosteric modulatory site. Brit J Pharmacol 137: 589-596. https://doi.org/10.1038/sj.bjp.0704829
|
| [117] |
Oz M, Zhang L, Morales M (2002) Endogenous cannabinoid, anandamide, acts as a noncompetitive inhibitor on 5-HT3 receptor-mediated responses in Xenopus oocytes. Synapse 46: 150-156. https://doi.org/10.1002/syn.10121
|
| [118] |
Xiong W, Koo BN, Morton R, et al. (2011) Psychotropic and nonpsychotropic cannabis derivatives inhibit human 5-HT3A receptors through a receptor desensitization-dependent mechanism. Neuroscience 184: 28-37. https://doi.org/10.1016/j.neuroscience.2011.03.066
|
| [119] |
Yang KH, Galadari S, Isaev D, et al. (2010) The nonpsychoactive cannabinoid cannabidiol inhibits 5-hydroxytryptamine3A receptor-mediated currents in Xenopus laevis oocytes. J Pharmacol Exp Ther 333: 547-554. https://doi.org/10.1124/jpet.109.162594
|
| [120] |
Yang KHS, Isaev D, Morales M, et al. (2010) The effect of Δ9-tetrahydrocannabinol on 5-HT3 receptors depends on the current density. Neuroscience 171: 40-49. https://doi.org/10.1016/j.neuroscience.2010.08.044
|
| [121] |
Fan P (1995) Cannabinoid agonists inhibit the activation of 5-HT3 receptors in rat nodose ganglion neurons. J Neurophysiol 73: 907-910. https://doi.org/10.1152/jn.1995.73.2.907
|
| [122] |
Xiong W, Hosoi M, Koo B-N, et al. (2008) Anandamide Inhibition of 5-HT<sub>3A</sub> Receptors Varies with Receptor Density and Desensitization. Mol Pharmacol 73: 314-322. https://doi.org/10.1124/mol.107.039149
|
| [123] |
Beaulieu JM, Espinoza S, Gainetdinov RR (2015) Dopamine receptors - IUPHAR Review 13. Br J Pharmacol 172: 1-23. https://doi.org/10.1111/bph.12906
|
| [124] |
Seeman P (2016) Cannabidiol is a partial agonist at dopamine D2High receptors, predicting its antipsychotic clinical dose. Transl Psychiatry 6: e920. https://doi.org/10.1038/tp.2016.195
|
| [125] |
Hansen KB, Wollmuth LP, Bowie D, et al. (2021) Structure, Function, and Pharmacology of Glutamate Receptor Ion Channels. Pharmacol Rev 73: 1469-1658. https://doi.org/10.1124/pharmrev.120.000131
|
| [126] |
Yu Y, Yang Z, Jin B, et al. (2020) Cannabidiol inhibits febrile seizure by modulating AMPA receptor kinetics through its interaction with the N-terminal domain of GluA1/GluA2. Pharmacol Res 161: 105128. https://doi.org/10.1016/j.phrs.2020.105128
|
| [127] |
Akinshola BE, Taylor RE, Ogunseitan AB, et al. (1999) Anandamide inhibition of recombinant AMPA receptor subunits in Xenopus oocytes is increased by forskolin and 8-bromo-cyclic AMP. N-S Arch Pharmacol 360: 242-248. https://doi.org/10.1007/s002109900078
|
| [128] |
Hampson AJ, Bornheim LM, Scanziani M, et al. (1998) Dual effects of anandamide on NMDA receptor-mediated responses and neurotransmission. J Neurochem 70: 671-676. https://doi.org/10.1046/j.1471-4159.1998.70020671.x
|
| [129] |
Malinowska B, Zakrzeska A, Kurz CM, et al. (2010) Involvement of central β2-adrenergic, NMDA and thromboxane A2 receptors in the pressor effect of anandamide in rats. N-S Arch Pharmacol 381: 349-360. https://doi.org/10.1007/s00210-010-0497-6
|
| [130] |
Sigel E, Steinmann ME (2012) Structure, Function, and Modulation of GABAA Receptors*. J Biol Chem 287: 40224-40231. https://doi.org/10.1074/jbc.R112.386664
|
| [131] |
Olsen RW (2018) GABAA receptor: Positive and negative allosteric modulators. Neuropharmacology 136: 10-22. https://doi.org/10.1016/j.neuropharm.2018.01.036
|
| [132] |
Koe BK, Milne GM, Weissman A, et al. (1985) Enhancement of brain [3H]flunitrazepam binding and analgesic activity of synthetic cannabimimetics. Eur J Pharmacol 109: 201-212. https://doi.org/10.1016/0014-2999(85)90421-2
|
| [133] | KOE BK, WEISSMAN A (1981) Facilitation of Benzodiazepine Binding by Levonantradol. J Clin Pharmacol 21: 397S-405S. https://doi.org/10.1002/j.1552-4604.1981.tb02619.x |
| [134] |
Bakas T, van Nieuwenhuijzen PS, Devenish SO, et al. (2017) The direct actions of cannabidiol and 2-arachidonoyl glycerol at GABA(A) receptors. Pharmacol Res 119: 358-370. https://doi.org/10.1016/j.phrs.2017.02.022
|
| [135] |
Anderson LL, Absalom NL, Abelev SV, et al. (2019) Coadministered cannabidiol and clobazam: Preclinical evidence for both pharmacodynamic and pharmacokinetic interactions. Epilepsia 60: 2224-2234. https://doi.org/10.1111/epi.16355
|
| [136] |
Ruffolo G, Cifelli P, Roseti C, et al. (2018) A novel GABAergic dysfunction in human Dravet syndrome. Epilepsia 59: 2106-2117. https://doi.org/10.1111/epi.14574
|
| [137] |
Yao L, Wells M, Wu X, et al. (2020) Membrane cholesterol dependence of cannabinoid modulation of glycine receptor. FASEB J 34: 10920-10930. https://doi.org/10.1096/fj.201903093R
|
| [138] |
Golovko T, Min R, Lozovaya N, et al. (2015) Control of Inhibition by the Direct Action of Cannabinoids on GABAA Receptors. Cereb Cortex 25: 2440-2455. https://doi.org/10.1093/cercor/bhu045
|
| [139] |
Voets T, Talavera K, Owsianik G, et al. (2005) Sensing with TRP channels. Nat Chem Biol 1: 85-92. https://doi.org/10.1038/nchembio0705-85
|
| [140] |
Caterina MJ (2014) TRP channel cannabinoid receptors in skin sensation, homeostasis, and inflammation. ACS Chem Neurosci 5: 1107-1116. https://doi.org/10.1021/cn5000919
|
| [141] |
Akopian AN, Ruparel NB, Jeske NA, et al. (2009) Role of ionotropic cannabinoid receptors in peripheral antinociception and antihyperalgesia. Trends Pharmacol Sci 30: 79-84. https://doi.org/10.1016/j.tips.2008.10.008
|
| [142] |
Di Marzo V, De Petrocellis L (2010) Endocannabinoids as regulators of transient receptor potential (TRP) channels: A further opportunity to develop new endocannabinoid-based therapeutic drugs. Curr Med Chem 17: 1430-1449. https://doi.org/10.2174/092986710790980078
|
| [143] |
De Petrocellis L, Ligresti A, Moriello AS, et al. (2011) Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Brit J Pharmacol 163: 1479-1494. https://doi.org/10.1111/j.1476-5381.2010.01166.x
|
| [144] |
De Petrocellis L, Orlando P, Moriello AS, et al. (2012) Cannabinoid actions at TRPV channels: effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol (Oxf) 204: 255-266. https://doi.org/10.1111/j.1748-1716.2011.02338.x
|
| [145] |
De Petrocellis L, Vellani V, Schiano-Moriello A, et al. (2008) Plant-derived cannabinoids modulate the activity of transient receptor potential channels of ankyrin type-1 and melastatin type-8. J Pharmacol Exp Ther 325: 1007-1015. https://doi.org/10.1124/jpet.107.134809
|
| [146] |
McKemy DD (2005) How cold is it? TRPM8 and TRPA1 in the molecular logic of cold sensation. Mol Pain 1: 16. https://doi.org/10.1186/1744-8069-1-16
|
| [147] |
De Petrocellis L, Schiano Moriello A, Imperatore R, et al. (2012) A re-evaluation of 9-HODE activity at TRPV1 channels in comparison with anandamide: enantioselectivity and effects at other TRP channels and in sensory neurons. Br J Pharmacol 167: 1643-1651. https://doi.org/10.1111/j.1476-5381.2012.02122.x
|
| [148] |
Heblinski M, Santiago M, Fletcher C, et al. (2020) Terpenoids Commonly Found in Cannabis sativa Do Not Modulate the Actions of Phytocannabinoids or Endocannabinoids on TRPA1 and TRPV1 Channels. Cannabis Cannabinoid Res 5: 305-317. https://doi.org/10.1089/can.2019.0099
|
| [149] |
De Petrocellis L, Starowicz K, Moriello AS, et al. (2007) Regulation of transient receptor potential channels of melastatin type 8 (TRPM8): effect of cAMP, cannabinoid CB(1) receptors and endovanilloids. Exp Cell Res 313: 1911-1920. https://doi.org/10.1016/j.yexcr.2007.01.008
|
| [150] |
Catterall WA (2012) Voltage-gated sodium channels at 60: structure, function and pathophysiology. J Physiol 590: 2577-2589. https://doi.org/10.1113/jphysiol.2011.224204
|
| [151] |
Ghovanloo MR, Aimar K, Ghadiry-Tavi R, et al. (2016) Physiology and Pathophysiology of Sodium Channel Inactivation. Curr Top Membr 78: 479-509. https://doi.org/10.1016/bs.ctm.2016.04.001
|
| [152] |
Ghovanloo MR, Ruben PC (2022) Cannabidiol and Sodium Channel Pharmacology: General Overview, Mechanism, and Clinical Implications. Neuroscientist 28: 318-334. https://doi.org/10.1177/10738584211017009
|
| [153] |
Catterall WA, Swanson TM (2015) Structural Basis for Pharmacology of Voltage-Gated Sodium and Calcium Channels. Mol Pharmacol 88: 141-150. https://doi.org/10.1124/mol.114.097659
|
| [154] |
Jiang D, Shi H, Tonggu L, et al. (2020) Structure of the Cardiac Sodium Channel. Cell 180: 122-134.e110. https://doi.org/10.1016/j.cell.2019.11.041
|
| [155] |
West JW, Patton DE, Scheuer T, et al. (1992) A cluster of hydrophobic amino acid residues required for fast Na(+)-channel inactivation. Proc Natl Acad Sci U S A 89: 10910-10914. https://doi.org/10.1073/pnas.89.22.10910
|
| [156] |
Ghovanloo MR, Shuart NG, Mezeyova J, et al. (2018) Inhibitory effects of cannabidiol on voltage-dependent sodium currents. J Biol Chem 293: 16546-16558. https://doi.org/10.1074/jbc.RA118.004929
|
| [157] |
Sait LG, Sula A, Ghovanloo M-R, et al. (2020) Cannabidiol interactions with voltage-gated sodium channels. eLife 9: e58593. https://doi.org/10.7554/eLife.58593
|
| [158] |
Johnson JR, Burnell-Nugent M, Lossignol D, et al. (2010) Multicenter, double-blind, randomized, placebo-controlled, parallel-group study of the efficacy, safety, and tolerability of THC:CBD extract and THC extract in patients with intractable cancer-related pain. J Pain Symptom Manage 39: 167-179. https://doi.org/10.1016/j.jpainsymman.2009.06.008
|
| [159] |
Turkanis SA, Partlow LM, Karler R (1991) Delta-9-tetrahydrocannabinol depresses inward sodium current in mouse neuroblastoma cells. Neuropharmacology 30: 73-77. https://doi.org/10.1016/0028-3908(91)90045-D
|
| [160] |
Kushner J, Ferrer X, Marx SO (2019) Roles and Regulation of Voltage-gated Calcium Channels in Arrhythmias. J Innov Card Rhythm Manag 10: 3874-3880. https://doi.org/10.19102/icrm.2019.101006
|
| [161] |
Ali RM, Al Kury LT, Yang KH, et al. (2015) Effects of cannabidiol on contractions and calcium signaling in rat ventricular myocytes. Cell Calcium 57: 290-299. https://doi.org/10.1016/j.ceca.2015.02.001
|
| [162] |
Oz M, Tchugunova Y, Dinc M (2004) Differential effects of endogenous and synthetic cannabinoids on voltage-dependent calcium fluxes in rabbit T-tubule membranes: comparison with fatty acids. Eur J Pharmacol 502: 47-58. https://doi.org/10.1016/j.ejphar.2004.08.052
|
| [163] |
Al Kury LT, Voitychuk OI, Yang KH, et al. (2014) Effects of the endogenous cannabinoid anandamide on voltage-dependent sodium and calcium channels in rat ventricular myocytes. Br J Pharmacol 171: 3485-3498. https://doi.org/10.1111/bph.12734
|
| [164] |
Chemin J, Monteil A, Perez-Reyes E, et al. (2001) Direct inhibition of T-type calcium channels by the endogenous cannabinoid anandamide. Embo J 20: 7033-7040. https://doi.org/10.1093/emboj/20.24.7033
|
| [165] | Hille B (2001) Ion channels of excitable membranes. Sunderland, Mass.: Sinauer Sunderland, Mass. |
| [166] |
McCoy MT, Jayanthi S, Cadet JL (2021) Potassium Channels and Their Potential Roles in Substance Use Disorders. Int J Mol Sci 22: 1249. https://doi.org/10.3390/ijms22031249
|
| [167] | Zhang HB, Heckman L, Niday Z, et al. (2022) Cannabidiol activates neuronal Kv7 channels. Elife 11. https://doi.org/10.7554/eLife.73246 |
| [168] |
Isaev D, Shabbir W, Dinc EY, et al. (2022) Cannabidiol Inhibits Multiple Ion Channels in Rabbit Ventricular Cardiomyocytes. Front Pharmacol 13. https://doi.org/10.3389/fphar.2022.821758
|
| [169] |
Topal L, Naveed M, Orvos P, et al. (2021) The electrophysiological effects of cannabidiol on action potentials and transmembrane potassium currents in rabbit and dog cardiac ventricular preparations. Arch Toxicol 95: 2497-2505. https://doi.org/10.1007/s00204-021-03086-0
|
| [170] |
Poling JS, Rogawski MA, Salem N, et al. (1996) Anandamide, an endogenous cannabinoid, inhibits Shaker-related voltage-gated K+ channels. Neuropharmacology 35: 983-991. https://doi.org/10.1016/0028-3908(96)00130-X
|
| [171] |
Bondarenko AI, Panasiuk O, Okhai I, et al. (2017) Direct activation of Ca(2+) and voltage-gated potassium channels of large conductance by anandamide in endothelial cells does not support the presence of endothelial atypical cannabinoid receptor. Eur J Pharmacol 805: 14-24. https://doi.org/10.1016/j.ejphar.2017.03.038
|
| [172] |
Amorós I, Barana A, Caballero R, et al. (2010) Endocannabinoids and cannabinoid analogues block human cardiac Kv4.3 channels in a receptor-independent manner. J Mol Cell Cardiol 48: 201-210. https://doi.org/10.1016/j.yjmcc.2009.07.011
|
| [173] |
Li Q, Ma HJ, Song SL, et al. (2012) Effects of anandamide on potassium channels in rat ventricular myocytes: a suppression of I(to) and augmentation of K(ATP) channels. Am J Physiol Cell Physiol 302: C924-930. https://doi.org/10.1152/ajpcell.00228.2011
|
| [174] |
Oz M, Yang KH, Dinc M, et al. (2007) The endogenous cannabinoid anandamide inhibits cromakalim-activated K+ currents in follicle-enclosed Xenopus oocytes. J Pharmacol Exp Ther 323: 547-554. https://doi.org/10.1124/jpet.107.125336
|
| [175] |
Oliver D, Lien CC, Soom M, et al. (2004) Functional conversion between A-type and delayed rectifier K+ channels by membrane lipids. Science 304: 265-270. https://doi.org/10.1126/science.1094113
|
| [176] |
Van den Bossche I, Vanheel B (2000) Influence of cannabinoids on the delayed rectifier in freshly dissociated smooth muscle cells of the rat aorta. Br J Pharmacol 131: 85-93. https://doi.org/10.1038/sj.bjp.0703521
|
| [177] |
Vignali M, Benfenati V, Caprini M, et al. (2009) The endocannabinoid anandamide inhibits potassium conductance in rat cortical astrocytes. Glia 57: 791-806. https://doi.org/10.1002/glia.20807
|
| [178] |
Maingret F, Patel AJ, Lazdunski M, et al. (2001) The endocannabinoid anandamide is a direct and selective blocker of the background K(+) channel TASK-1. Embo J 20: 47-54. https://doi.org/10.1093/emboj/20.1.47
|
| [179] |
Gantz SC, Bean BP (2017) Cell-Autonomous Excitation of Midbrain Dopamine Neurons by Endocannabinoid-Dependent Lipid Signaling. Neuron 93: 1375-1387.e1372. https://doi.org/10.1016/j.neuron.2017.02.025
|
| [180] |
Spivak CE, Kim W, Liu Q-R, et al. (2012) Blockade of β-cell KATP channels by the endocannabinoid, 2-arachidonoylglycerol. Biochem Bioph Res Co 423: 13-18. https://doi.org/10.1016/j.bbrc.2012.05.042
|
Figures(1) / Tables(1)

Nicholas J. D. Wright. A review of the direct targets of the cannabinoids cannabidiol, Δ9-tetrahydrocannabinol, N-arachidonoylethanolamine and 2-arachidonoylglycerol[J]. AIMS Neuroscience, 2024, 11(2): 144-165. doi: 10.3934/Neuroscience.2024009







 DownLoad:
DownLoad: