Citation: Marios G. Krokidis. Identification of biomarkers associated with Parkinson’s disease by gene expression profiling studies and bioinformatics analysis[J]. AIMS Neuroscience, 2019, 6(4): 333-345. doi: 10.3934/Neuroscience.2019.4.333

| [1] |
Obeso J, Rodriguez-Oroz M, Goetz C, et al. (2010) Missing pieces in the Parkinson's disease puzzle. Nat Med 16: 653–661. doi: 10.1038/nm.2165

|
| [2] | Musgrove R, Helwig M, Bae E, et al. (2019) Oxidative stress in vagal neurons promotes parkinsonian pathology and intercellular α-synuclein transfer. J Clin Invest 130: 3738–3753. |
| [3] |
Lill C (2016) Genetics of Parkinson's disease. Mol Cell Probes 30: 386–396. doi: 10.1016/j.mcp.2016.11.001
|
| [4] |
Reale M, Iarlori C, Thomas A, et al. (2009) Peripheral cytokines profile in Parkinson's disease. Brain Behav Immun 23: 55–63. doi: 10.1016/j.bbi.2008.07.003
|
| [5] |
Dias V, Junn E, Mouradian M (2013) The role of oxidative stress in Parkinson's disease. J Parkinsons Dis 3: 461–491. doi: 10.3233/JPD-130230
|
| [6] |
Taylor J, Main B, Crack P (2013) Neuroinflammation and oxidative stress: co-conspirators in the pathology of Parkinson's disease. Neurochem Int 62: 803–819. doi: 10.1016/j.neuint.2012.12.016
|
| [7] | Kalia L, Lang A (2015) Parkinson's disease. Lancet 386 :896–912. |
| [8] | Valdes P, Schneider B (2016) Gene therapy: A promising approach for neuroprotection in Parkinson's disease? Front Neuroanat 10: 123. |
| [9] |
Ferrer I, Lopez-Gonzalez I, Carmona M, et al. (2012) Neurochemistry and the non-motor aspects of PD. Neurobiol Dis 46: 508–526. doi: 10.1016/j.nbd.2011.10.019
|
| [10] |
Scherzer C, Grass J, Liao Z, et al. (2008) GATA transcription factors directly regulate the Parkinson's disease-linked gene α-synuclein. Proc Natl Acad Sci USA 105: 10907–10912. doi: 10.1073/pnas.0802437105
|
| [11] |
Miller D, O'Callaghan J (2015) Biomarkers of Parkinson's disease: Present and future. Metabolism 64: S40–S46. doi: 10.1016/j.metabol.2014.10.030
|
| [12] |
Harraz M, Dawson T, Dawson V (2011) MicroRNAs in Parkinson's disease. J Chem Neuroanat 42: 127–130. doi: 10.1016/j.jchemneu.2011.01.005
|
| [13] | Gasser T (2009) Genomic and proteomic biomarkers for Parkinson disease. Neurology 72: 27–31. |
| [14] |
Haque M, Thomas K, D'Souza C, et al. (2008) Cytoplasmic Pink1 activity protects neurons from dopaminergic neurotoxin MPTP. Proc Natl Acad Sci USA 105: 1716–1721. doi: 10.1073/pnas.0705363105
|
| [15] |
Valente E, Bentivoglio A, Dixon P, et al. (2001) Localization of a novel locus for autosomal recessive early-onset parkinsonism, PARK6, on human chromosome 1p35–p36. Am J Hum Genet 68: 895–900. doi: 10.1086/319522
|
| [16] |
Kitada T, Asakawa S, Hattori N, et al. (1998) Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 392: 605–608. doi: 10.1038/33416
|
| [17] | McKeith I (2004) Dementia with Lewy bodies. Dialogues Clin Neurosci 3: 333–341. |
| [18] | Farrer M (2006) Genetics of Parkinson disease: paradigm shifts and future prospects. Nat Rev Genet 7: 306–318. |
| [19] |
Funayama M, Hasegawa K, Kowa H, et al. (2002) A new locus for Parkinson's disease (PARK8) maps to chromosome 12p11.2–q13.1. Ann Neurol 51: 296–301. doi: 10.1002/ana.10113
|
| [20] |
Mencacci N, Isaias I, Reich M, et al. (2014) Parkinson's disease in GTP cyclohydrolase 1 mutation carriers. Brain 137: 2480–2492. doi: 10.1093/brain/awu179
|
| [21] |
Olgiati S, Quadri M, Fang M, et al. (2016) DNAJC6 mutations associated with early-onset Parkinson's disease. Ann Neurol 79: 244–256. doi: 10.1002/ana.24553
|
| [22] |
Jiang F, Wu Q, Sun S, et al. (2019) Identification of potential diagnostic biomarkers for Parkinson's disease. FEBS Open Bio 9: 1460–1468. doi: 10.1002/2211-5463.12687
|
| [23] |
Wang Q, Zhang Y, Wang M, et al. (2019) The landscape of multiscale transcriptomic networks and key regulators in Parkinson's disease. Nat Commun 10: 5234. doi: 10.1038/s41467-019-13144-y
|
| [24] |
Chi J, Xie Q, Jia J, et al. (2018) Integrated analysis and identification of novel biomarkers in Parkinson's disease. Front Aging Neurosci 10: 178. doi: 10.3389/fnagi.2018.00178
|
| [25] |
Kong P, Lei P, Zhang S, et al. (2018) Integrated microarray analysis provided a new insight of the pathogenesis of Parkinson's disease. Neurosci Lett 662: 51–58. doi: 10.1016/j.neulet.2017.09.051
|
| [26] |
Wang C, Chen L, Yang Y, et al. (2019) Identification of potential blood biomarkers for Parkinson's disease by gene expression and DNA methylation data integration analysis. Clin Epigenetics 11: 24. doi: 10.1186/s13148-019-0621-5
|
| [27] |
George G, Singh S, Lokappa S, et al. (2019). Gene co-expression network analysis for identifying genetic markers in Parkinson's disease-a three-way comparative approach. Genomics 111: 819–830. doi: 10.1016/j.ygeno.2018.05.005
|
| [28] |
Kim J, Inoue K, Ishii J, et al. (2007) A MicroRNA feedback circuit in midbrain dopamine neurons. Science 317: 1220–1224. doi: 10.1126/science.1140481
|
| [29] |
Wang G, van der Walt J, Mayhew G, et al. (2008) Variation in the miRNA-433 binding site of FGF20 confers risk for Parkinson disease by overexpression of alpha-synuclein. Am J Hum Genet 82: 283–289. doi: 10.1016/j.ajhg.2007.09.021
|
| [30] |
Mouradian M (2012) MicroRNAs in Parkinson's disease. Neurobiol Dis 46: 279–284. doi: 10.1016/j.nbd.2011.12.046
|
| [31] |
Junn E, Lee K, Jeong B, et al. (2009) Mouradian Repression of alpha-synuclein expression and toxicity by microRNA-7. Proc Natl Acad Sci USA 106: 13052–13057. doi: 10.1073/pnas.0906277106
|
| [32] |
Doxakis E (2010) Post-transcriptional regulation of alpha-synuclein expression by mir-7 and mir-153. Biol Chem 285: 12726–12734. doi: 10.1074/jbc.M109.086827
|
| [33] |
van der Walt J, Noureddine M, Kittappa R, et al. (2004) Fibroblast growth factor 20 polymorphisms and haplotypes strongly influence risk of Parkinson disease. Am J Hum Genet 74: 1121–1127. doi: 10.1086/421052
|
| [34] |
Schulz J, Takousis P, Wohlers I, et al. (2019) Meta‐analyses identify differentially expressed microRNAs in Parkinson's disease. Ann Neurol 85: 835–851. doi: 10.1002/ana.25490
|
| [35] |
Uwatoko H, Hama Y, Iwata I, et al. (2019) Identification of plasma microRNA expression changes in multiple system atrophy and Parkinson's disease. Mol Brain 12: 49. doi: 10.1186/s13041-019-0471-2
|
| [36] |
Chen L, Yang J, Lü J, et al. (2018) Identification of aberrant circulating miRNAs in Parkinson's disease plasma samples. Brain Behav 8: e00941. doi: 10.1002/brb3.941
|
| [37] | Goh S, Chao Y, Dheen S, et al. (2019) Role of MicroRNAs in Parkinson's Disease. Int J Mol Sci 20: pii: E5649. |
| [38] | Peeraully T, Tan E (2012) Genetic variants in sporadic Parkinson's disease: East vs West. Parkinsonism Relat Disord 18: S63–S65. |
| [39] |
Chikina M, Gerald C, Li X, et al. (2015) Low-variance RNAs identify Parkinson's disease molecular signature in blood. Mov Disord 30: 813–821. doi: 10.1002/mds.26205
|
| [40] |
Qian Y, Yang X, Xu S, et al. (2018) Detection of microbial 16S rRNA gene in the blood of patients with Parkinson's disease. Front Aging Neurosci 10: 156. doi: 10.3389/fnagi.2018.00156
|
| [41] |
Li Y, Wong G, Humphrey J, et al. (2019) Prioritizing Parkinson's disease genes using population-scale transcriptomic data. Nat Commun 10: 994. doi: 10.1038/s41467-019-08912-9
|
| [42] |
Chang D, Nalls M, Hallgrímsdóttir I, et al. (2017) A meta-analysis of genome-wide association studies identifies 17 new Parkinson's disease risk loci. Nat Genet 49: 1511–1516. doi: 10.1038/ng.3955
|
| [43] |
Garnier S, Truong V, Brocheton J, et al. (2013) Genome-wide haplotype analysis of cis expression quantitative trait loci in monocytes. PLoS Genet 9: e1003240. doi: 10.1371/journal.pgen.1003240
|
| [44] |
Sekiyama K, Takamatsu Y, Waragai M, et al. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226–235. doi: 10.1016/j.bbrc.2014.06.028
|
| [45] | Servant N, Roméjon J, Gestraud P, et al. (2014) Bioinformatics for precision medicine in oncology: principles and application to the SHIVA clinical trial. Front Genet 5: 152. |
| [46] |
Dong N, Zhang X, Liu Q (2017) Identification of therapeutic targets for Parkinson's disease via bioinformatics analysis. Mol Med Rep 15: 731–735. doi: 10.3892/mmr.2016.6044
|
| [47] | Al-Ouran R, Wan Y, Mangleburg C, et al. (2019) A portal to visualize transcriptome profiles in mouse models of neurological disorders. Genes (Basel)10: pii: E759. |
| [48] |
Fu L, Fu K (2015) Analysis of Parkinson's disease pathophysiology using an integrated genomics-bioinformatics approach. Pathophysiology 22: 15–29. doi: 10.1016/j.pathophys.2014.10.002
|
| [49] |
Mariani E, Frabetti F, Tarozzi A, et al. (2016) Meta-Analysis of Parkinson's Disease transcriptome data using TRAM software: Whole substantia nigra tissue and single dopamine neuron differential gene expression. PLoS One 11: e0161567. doi: 10.1371/journal.pone.0161567
|
| [50] |
Glaab E (2018) Computational systems biology approaches for Parkinson's disease. Cell Tissue Res 373: 91–109. doi: 10.1007/s00441-017-2734-5
|
| [51] |
Lottaz C, Toedling J, Spang R (2007) Annotation-based distance measures for patient subgroup discovery in clinical microarray studies. Bioinformatics 23: 2256–2264. doi: 10.1093/bioinformatics/btm322
|
| [52] |
Hu Y, Pan Z, Hu Y, et al. (2017) Network and pathway-based analyses of genes associated with Parkinson's disease. Mol Neurobiol 54: 4452–4465. doi: 10.1007/s12035-016-9998-8
|
| [53] |
Cheng HC, Ulane C, Burke R (2010) Clinical progression in Parkinson disease and the neurobiology of axons. Ann Neurol 67: 715–725. doi: 10.1002/ana.21995
|
| [54] |
Palfi S, Gurruchaga J, Ralph G, et al. (2014) Long-term safety and tolerability of ProSavin, a lentiviral vector-based gene therapy for Parkinson's disease: A dose escalation, open-label, phase 1/2 trial. Lancet 383: 1138–1146. doi: 10.1016/S0140-6736(13)61939-X
|
| [55] |
Diao H, Li X, Hu S, et al. (2012) Gene expression profiling combined with bioinformatics analysis identify biomarkers for Parkinson disease. PLoS One 7: e52319. doi: 10.1371/journal.pone.0052319
|
| [56] | Xu C, Chen J, Xu X, et al. (2018) Potential therapeutic drugs for Parkinson's disease based on data mining and bioinformatics analysis. Parkinsons Dis 2018: 3464578. |
| [57] | Airavaara M, Voutilainen MH, Wang Y, et al. (2012) Neurorestoration. Parkinsonism Relat Disord 18: S143–146. |
| [58] |
Francardo V, Schmitz Y, Sulzer D, et al. (2017) Neuroprotection and neurorestoration as experimental therapeutics for Parkinson's disease. Exp Neurol 298: 137–147. doi: 10.1016/j.expneurol.2017.10.001
|
| [59] | Nayak L, Henchcliffe C (2008) Rasagiline in treatment of Parkinson's disease. Neuropsychiatr Dis Treat 4: 23–32. |
| [60] |
Marsili L, Marconi R, Colosimo C (2017) Treatment strategies in early Parkinson's disease. Int Rev Neurobiol 132: 345–360. doi: 10.1016/bs.irn.2017.01.002
|
| [61] |
Magyar K, Stocchi F, Fossati C, et al. (2015) Rasagiline for the treatment of Parkinson's disease: an update. Expert Opin Pharmacother 16: 2231–2241. doi: 10.1517/14656566.2015.1086748
|
| [62] |
McCormack P (2014) Rasagiline: A review of its use in the treatment of idiopathic Parkinson's disease. CNS Drugs 28: 1083–1097. doi: 10.1007/s40263-014-0206-y
|
| [63] |
Alborghetti M, Nicoletti F (2019) Different generations of Type-B monoamine oxidase inhibitors in Parkinson's disease: From bench to bedside. Curr Neuropharmacol 17: 861–873. doi: 10.2174/1570159X16666180830100754
|
| [64] |
Bette S, Shpiner D, Singer C, et al. (2018) Safinamide in the management of patients with Parkinson's disease not stabilized on levodopa: A review of the current clinical evidence. Ther Clin Risk Manag 14: 1737–1745. doi: 10.2147/TCRM.S139545
|
| [65] |
Leegwater-Kim J, Waters C (2006) Tolcapone in the management of Parkinson's disease. Expert Opin Pharmacother 7: 2263–2270. doi: 10.1517/14656566.7.16.2263
|
| [66] | Jiang D, Wang H, Wang Y, et al. (2019) Rasagiline combined with levodopa therapy versus levodopa monotherapy for patients with Parkinson's disease: A systematic review. Neurol Sci, 1–9. |
| [67] |
Avila A, Caballol N, Martín-Baranera M, et al. (2019) Rasagiline and safinamide as a dopamine-sparing therapy for Parkinson's disease. Acta Neurol Scand 140: 23–31. doi: 10.1111/ane.13096
|
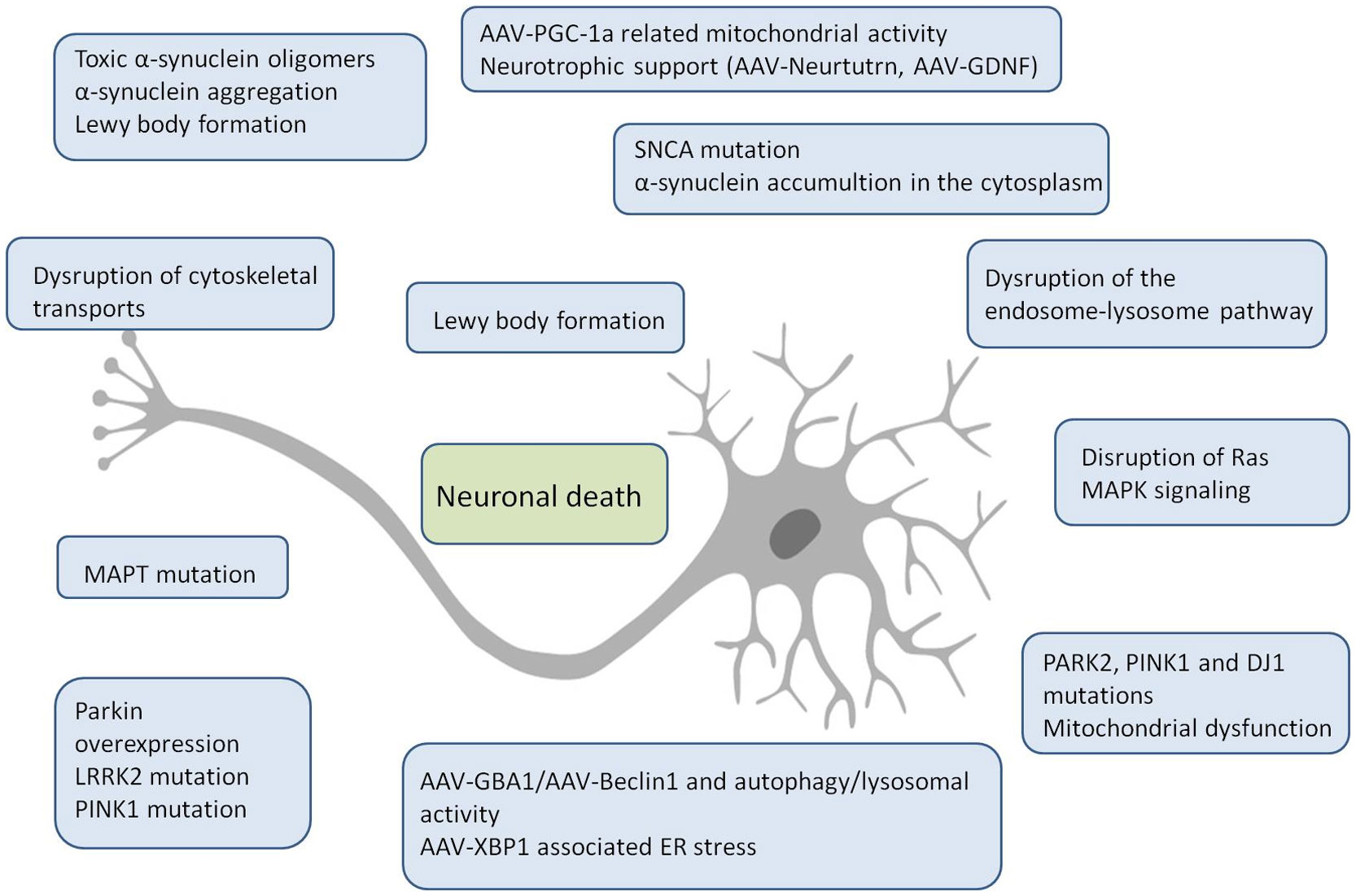
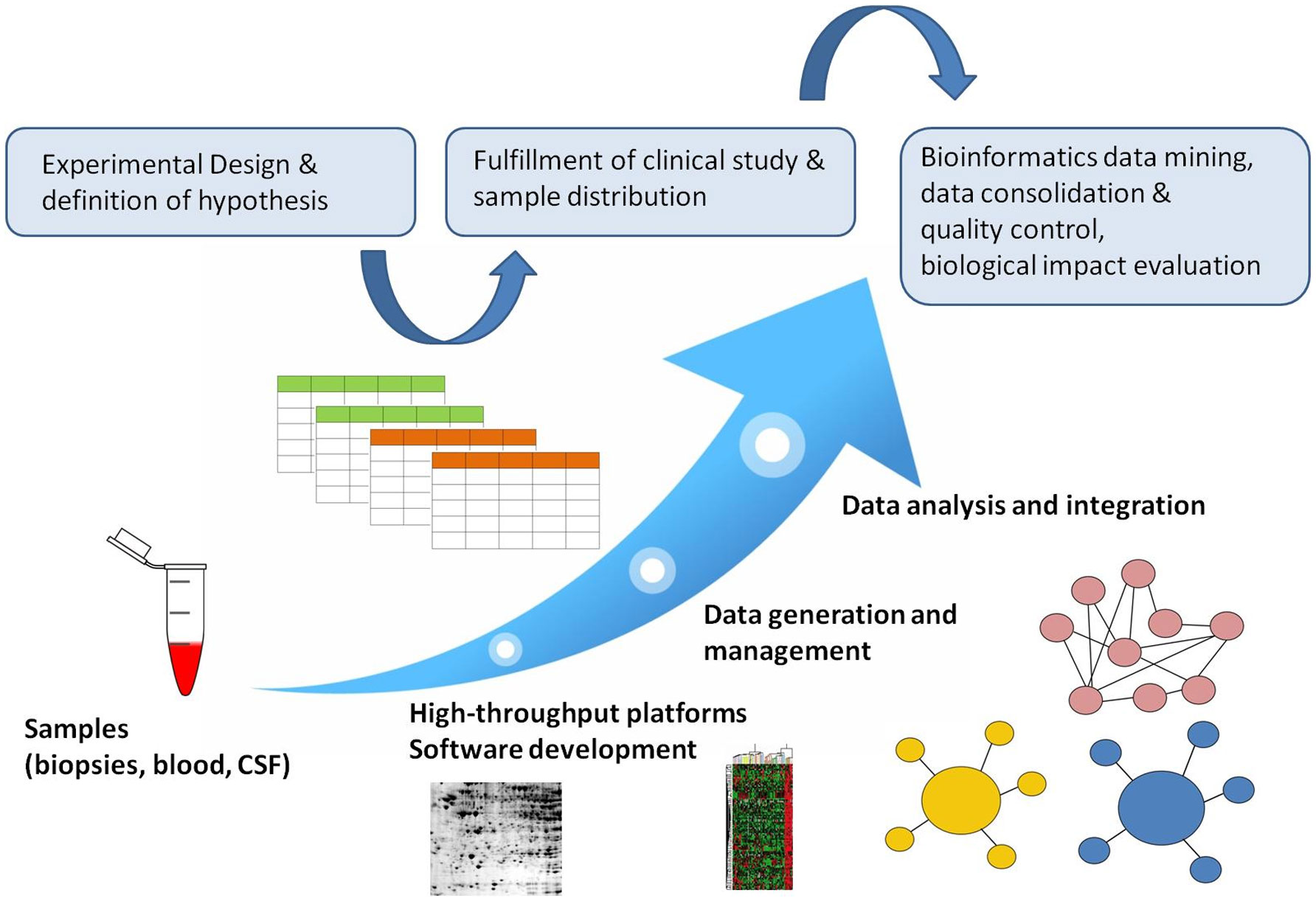
Figures(2)

Marios G. Krokidis. Identification of biomarkers associated with Parkinson’s disease by gene expression profiling studies and bioinformatics analysis[J]. AIMS Neuroscience, 2019, 6(4): 333-345. doi: 10.3934/Neuroscience.2019.4.333







 DownLoad:
DownLoad: