Citation: Pranav Chintamani Joshi, Sugato Benerjee. Effects of glucocorticoids in depression: Role of astrocytes[J]. AIMS Neuroscience, 2018, 5(3): 200-210. doi: 10.3934/Neuroscience.2018.3.200
| [1] | Laaziz Abderrahim, El Mostafi Hicham, Elhessni Aboubaker, Azeroil Fatima, Touil Tarik, Boumlah Soufiane, Mesfioui Abdelhalim . Sex differences in behavioral, cognitive and voluntary ethanol-intake effects in Dexamethasone-induced depression-like state in Wistar rat. AIMS Neuroscience, 2022, 9(2): 228-249. doi: 10.3934/Neuroscience.2022012 |
| [2] | Ahlem Matallah, Rabie Guezi, Abdelmadjid Bairi . Repeated restraint stress induced neurobehavioral and sexual hormone disorders in male rats. AIMS Neuroscience, 2022, 9(2): 264-276. doi: 10.3934/Neuroscience.2022014 |
| [3] | Martina Valencia, Odra Santander, Eloísa Torres, Natali Zamora, Fernanda Muñoz, Rodrigo Pascual . Environmental enrichment reverses cerebellar impairments caused by prenatal exposure to a synthetic glucocorticoid. AIMS Neuroscience, 2022, 9(3): 320-344. doi: 10.3934/Neuroscience.2022018 |
| [4] | Fiammetta Iannuzzo, Federica Donia, Lorenzo Bette, Fabrizio Turiaco, Antonio Bruno . Neuromodulation techniques in combination for a short-intensive treatment of depression and anxiety: a case report. AIMS Neuroscience, 2025, 12(2): 140-146. doi: 10.3934/Neuroscience.2025009 |
| [5] | Dirk Roosterman, Graeme S. Cottrell . Astrocytes and neurons communicate via a monocarboxylic acid shuttle. AIMS Neuroscience, 2020, 7(2): 94-106. doi: 10.3934/Neuroscience.2020007 |
| [6] | Amanda G Bertollo, Roberta E Grolli, Marcos E Plissari, Vanessa A Gasparin, João Quevedo, Gislaine Z Réus, Margarete D Bagatini, Zuleide M Ignácio . Stress and serum cortisol levels in major depressive disorder: a cross-sectional study. AIMS Neuroscience, 2020, 7(4): 459-469. doi: 10.3934/Neuroscience.2020028 |
| [7] | Ashok Chakraborty, Anil Diwan . Depression and Parkinson's disease: a Chicken-Egg story. AIMS Neuroscience, 2022, 9(4): 479-490. doi: 10.3934/Neuroscience.2022027 |
| [8] | Ebtesam M Abd-El-Basset, Muddanna Sakkattu Rao, Solaiman M Alshawaf, Hasan Kh Ashkanani, Abdulaziz H Kabli . Tumor necrosis factor (TNF) induces astrogliosis, microgliosis and promotes survival of cortical neurons. AIMS Neuroscience, 2021, 8(4): 558-584. doi: 10.3934/Neuroscience.2021031 |
| [9] | Shahzaib Maqbool, Arham Ihtesham, Muhammad Nadeem Langove, Sara Jamal, Tabdar Jamal, Hafiz Abu Safian . Neuro-dermatological association between psoriasis and depression: an immune-mediated inflammatory process validating skin-brain axis theory. AIMS Neuroscience, 2021, 8(3): 340-354. doi: 10.3934/Neuroscience.2021018 |
| [10] | Nao Fukuwada, Miki Kanno, Satomi Yoshida, Kenjiro Seki . Gαq protein signaling in the bed nucleus of the stria terminalis regulate the lipopolysaccharide-induced despair-like behavior in mice. AIMS Neuroscience, 2020, 7(4): 438-458. doi: 10.3934/Neuroscience.2020027 |
Astrocytes or astroglia are highly heterogeneous glial cells that populate the brain and spinal cord, and are responsible for homeostasis of the central nervous system (CNS). Earlier, it was reported that, about 90% of the cortical tissue volume is made up of astrocytes, whereas the remaining 10% consists of neuronal cell bodies and blood vessels in the rat cerebral cortex [1]. However, the cellular composition of mammalian brain showed that astrocytes are only 20% of its glial cells; the majority (75%) are oligodendrocytes, while microglia amount to only 5% of glial cells in the grey matter [2]. Astrocytes, representing a large glial population in the mammalian brain [3], play a pivotal role in contributing to the brain metabolism [4] and are involved in glutamate clearance from the synapse and cycling of glutamine back into neurons (e.g. Glutamate-glutamine metabolism and transport) [5]. Astrocytes also release gliotransmitters such as glutamate and ATP [6], both onto neurons and other glial cells [6]. In contrast to chemical coupling at the synapse, astrocytes have extensive coupling via gap junctions, forming dynamic networks that passage molecules between astrocytes as well as to other cells in the CNS [7]. This passage includes interactions with the blood-brain barrier (BBB), mainly the vascular endothelial cells, as well as transporting metabolites to supply the energy needs of neurons [7,8].
The adrenal gland which secretes Glucocorticoids (GCs), in response to the signal from the hypothalamus, is responsible for producing a range of effects in response to stress. The release of GCs, a class of steroid hormones, is regulated by the hypothalamic-pituitary-adrenal (HPA) axis [9]. The HPA axis that belongs to neuroendocrine systems is commonly associated with stress signaling and the “fight-or-flight” response [10]. GCs act as potent transcriptional regulators that signal through two types of receptors: The high-affinity mineralocorticoid receptors (MRs) and the lower affinity glucocorticoid receptors (GRs) [11].
The MRs are associated with mechanisms involving lower GC concentrations, while the GRs respond to higher concentrations of GCs, such as levels associated with stressful experiences [12]. Alterations in the GC signaling are thought to contribute to disorders like Major Depression (MD) [13], Alzheimer's disease [14], and Cushing's syndrome [15], etc. A recent study on MD, showed that reduced levels of glial fibrillary acidic protein (GFAP) immunoreactive astrocytes [16] while the release of GCs in response to chronic stress leading to MD has been widely reported [17]. However, the role of stress hormones on astrocytic gene expression patterns, remain largely uncharacterized. Astrocytes are involved in primary biochemical processes and are one of the primary cell types in the brain [18]. An attempt is being made to explore the role of astrocytes in neuropsychiatric disorder like depression [19]. In this review, we primarily focus on the effects of the stress-induced GCs on the astrocytes and their implications in depression.
Initially, depression was viewed as a neuron-based disorder. This “neuron-centric” view of depression is at least partially supported by the known actions of commonly used antidepressants that modulate levels of the neurotransmitters in the brain (e.g. Serotonin by SSRIs) [20]. However, antidepressants target proteins expressed in multiple brain cell types (e.g. astrocytes express NMDA receptors and serotonin receptors [21]), thus distinct mechanisms and cells involved in antidepressant action remain largely unknown. A series of studies in the late 1990s and early 2000s made unexpected associations between depression and changes in cell density of specific cell types in the brain. Rajkowska and colleagues used cell counting approach to investigate how depression impacted cell density and morphology across various brain regions. Based on previous models, they suspected that depression would lead to cell death and decreased cell density in brain regions associated with mood disorders. They found that there were indeed decreases in cell density in areas such as the prefrontal cortex (PFC) [22] and hippocampus [23]. But, their findings were surprising regarding the cell type; the decreases in cell density were associated with morphologies consistent with both neurons and glial cells [24].
Psychiatric disorders like schizophrenia, bipolar disorder and MD involve reduced astrocytes and astroglial atrophy [25]. Postmortem studies of depressed patients showed reduced glial cell densities in the PFC, amygdala and hippocampus [24]. Banasr and Duman demonstrated that the depressive behavior can be induced by chemical astrocytes ablation in the PFC of rat [26]. Moreover, hippocampal astrocytic loss is associated with chronic stress [27] and also chronic stress interferes with glial cell metabolism via glutamatergic mechanisms [28]. It remains unclear as to how dysregulated GCs may be involved in the loss of astrocytes after stress.
These findings inspired biochemical studies that explored specific depression-linked decrease in astrocyte density based on reduced protein expression of GFAP (a biomarker for mature astrocytes). GFAP has shown its significant role in pathogenesis of various CNS pathologies [29]. The GFAP expression studies, showed a 15% loss of total astrocyte volume and decrease in expression in the brains of depressed individuals [24,30], while expression in patients increased after treatment with antidepressants. A different investigation on postmortem amygdale tissue from depressed patients showed reduction in nissl positive astrocytes [31]. Another astroglial marker, the calcium binding protein S100β was reported to be a marker of minor depression [32]. S100β was found to be reduced in the ventral PFC of depressed suicide victims (Figure 1A). The screening of 36 biological markers in 30 inbred mouse strains [33] showed that GFAP, S100β , glyoxalase 1 and histone deacetylase 5 responded to chronic treatment with fluoxetine. Similarly, reduction in the astroglial GFAP-positive cells and its overall immunoreactivity were detected in several animal models of chronic stress [24]. In situ hybridization and RT-qPCR gene expression studies with human brain samples allowed the detection and quantification of the mRNA present in brain sections and showed alterations in several transcripts for astrocyte proteins with depression in the locus coeruleus [24,34]. They include astrocytic markers like S100β, GFAP (Figure 1B), gap junction proteins (gap junction protein alpha 1 and 6 (Gja1 and Gja6)), and membrane channels proteins (Aquaporin-4 (AQP4)). It was also found that the reduction of glutamate transporter (GLT-1), glutamate aspartate transporter (GLAST), glutamine synthetase (GS), connexins (Cx43 and Cx30) [35,36] (Figure 1C). Similarly, the reports show a decrease in the mRNA expression of GLT-1, GLAST, and GS in the anterior cingulate and dorsolateral PFC [37], while some of these transcripts were reduced in other brain areas. This suggests that the astroglial cells may contribute towards altered neuroglial networks in different forms of depression.
In animal models, selective ablation of astroglial cells (L-a-aminoadipic acid, which envenoms astrocytes) triggered depressive behavior [24]. Pharmacological inhibition of astroglial gap junction connectivity [38] or astroglial plasmalemma glutamate transporters resulted in anhedonia [39] (Figure 1F), one of the key symptoms of depression. All these findings in animals subjected to depression models indicate astrocytic abnormality. After chronic exposure to chronic unpredictable stress (CUS), it was found an increase in glutamate release and reduced uptake in the hippocampus [40]. In the learned helplessness model, a similar trend of reduced glutamate uptake was also observed in the PFC, striatum and hippocampus [41]. Studies in rats also showed blockage of astroglial glutamate uptake and was sufficient to induce anhedonic state marked by decreased sucrose consumption [42]. Ketamine and riluzole, which acts as glutamate modulators, have shown antidepressant effects in patients and animal models [43].
Concurrent pathophysiology showed aberrant glutamatergic neurotransmission as a primary mechanism for major psychiatric disorders, including MD [44]. Astrocytes are fundamental elements in glutamatergic and GABAergic neurotransmission being the hubs for glutamate—glutamine and glutamine—GABA shuttles [45]. In the brains of MD patients, expression of astrocyte-specific glutamate transporters GLT-1, GLAST as well as glutamine synthase (GS) (Figure 1D, E) are reduced [24], indicating compromised astrocytic uptake of glutamate, as well as decreased glutamine production.
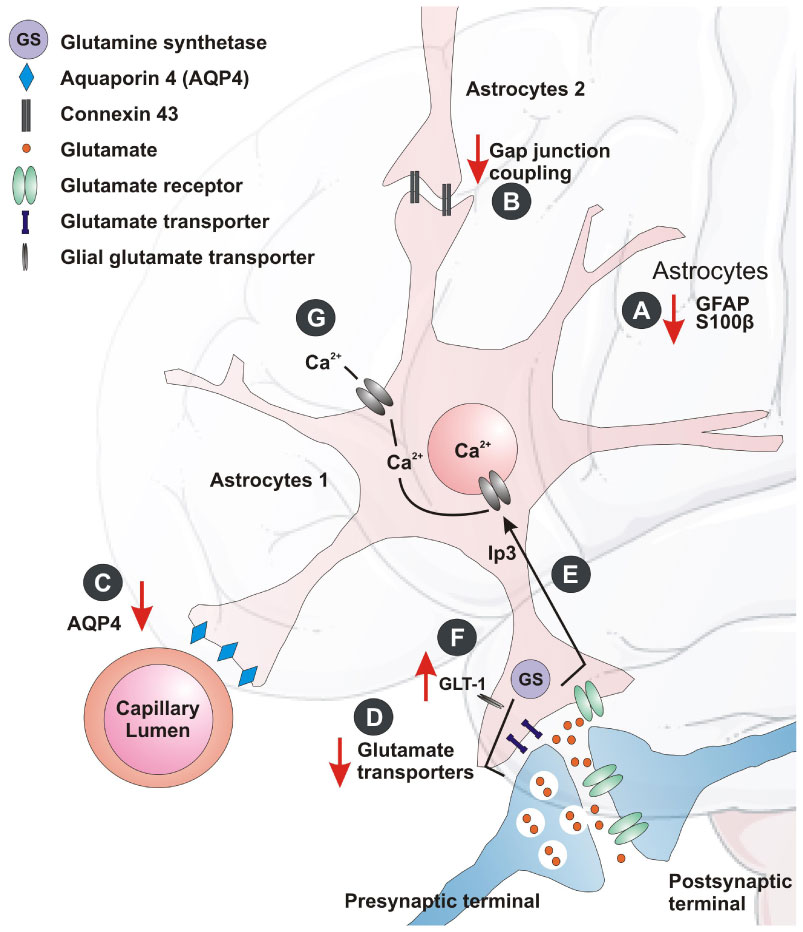 Figure 1. Astrocytic changes in the depressed brain. The crosstalk between the astrocytes and neurons is mediated at the tripartite synapse: A) Astrocyte marker GFAP and cytoplasmic protein S100β expression may alter with depression. B) Astrocyte-specific gap junction protein (Cx43) may alter during stress and depression. C) The water channel marker, AQP4 protein levels is found to reduce in depression. D and E) The expression of certain astrocyte glutamate transporters, as well as glutamine synthase may reduce in MD patients. F) There may be up-regulation of the hippocampal glial glutamate transporter (GLT-1) after chronic stress. G) Astrocytes may also coordinate their function through Ca2+ excitability and its downstream signaling in depression.
Figure 1. Astrocytic changes in the depressed brain. The crosstalk between the astrocytes and neurons is mediated at the tripartite synapse: A) Astrocyte marker GFAP and cytoplasmic protein S100β expression may alter with depression. B) Astrocyte-specific gap junction protein (Cx43) may alter during stress and depression. C) The water channel marker, AQP4 protein levels is found to reduce in depression. D and E) The expression of certain astrocyte glutamate transporters, as well as glutamine synthase may reduce in MD patients. F) There may be up-regulation of the hippocampal glial glutamate transporter (GLT-1) after chronic stress. G) Astrocytes may also coordinate their function through Ca2+ excitability and its downstream signaling in depression.Interactions between glial and neuronal cells may be impaired at synaptic level in depression-like conditions. Purines may modulate glia-neuron communication bi-directionally, restoring the synaptic efficacy and reversing depression-like behavior [46]. Another possible trigger for depressive episodes was the cytokine signaling in astrocytes [47]. Recent research focused on an alternative form of neuro-glial signaling, namely, secreted extracellular vesicles (EVS) [48]. The EVS, such as ectosomes and exosomes, capable of carrying mRNA and microRNAs (miRNA) have shown a role in intercellular signaling, and may also underlie depressive behavior. The transfer of exosomes to neurons is mediated through oligodendrocytes, microglia and astrocytes, which may either support neurons or promulgate the disease [48]. Thus neuroglia communication at the synaptic level is far from clear and warrants further research.
The mechanism underlying antidepressants action of fluoxetin is mediated by regulating astrocytic AQP4 levels that affects astrocyte morphology and further restore the functional glia-vasculature interface [48]. The decreased astrocyte-specific Cx43 levels, as stated earlier, appear to be related to antidepressant and anxiolytic phenotypes [49]. In accordance, the antidepressants show an intricate pattern linking astrocytes and connexins to address the mechanisms of action of these compounds [50]. Thus the role of astrocytes in depression is far from clear and needs further work.
The postmortem studies on humans and animals showed reduction in astrocyte density and function in the limbic regions of the brain suggest probable mechanism contributing to pathology of stress and GCs overproduction [51]. Indeed, a selective volume reduction of hippocampus and PFC was observed following chronic stress [51]. The neuronal proliferation in the dentate gyrus (DG) [52] was decreased after corticosterone treatment and psychosocial stress while the postmortem studies in patients with a history of high-dose steroid treatment. However, there was no reduction in the number of neurons [53]. This suggests that the volume reduction cannot be entirely due to stress-induced reduction of neurogenesis. The increased GCs immunoreactivity was observed in astrocytes of amygdala in depressed patients compared to healthy controls or bipolar disorder patients [54], suggesting that astrocytes may respond to changes in stress hormone levels.
Studies on chronic stress may result in reduced gliogenesis in the hippocampus and PFC [23,28] while in-vitro studies showed the dexamethasone (a synthetic glucocorticoid) blocking astrogliogenesis from neural precursor cells. These findings suggest that, the perturbed astroglial cells in stress is likely to contribute in region-specific volume changes commonly observed in stress-related pathologies [55].
The role of astrocytes in neurodegenerative diseases and inflammatory processes is well documented [56]. It is well reported that human postmortem tissue in major depressive disorders (MDD) have shown alterations in the expression of mRNA and protein for astrocyte markers such as GFAP, Cx40, Cx43, AQP4, S100β and glutamatergic markers including GLT-1, GLAST, and GS [53]. When the brain gets injured, astrocytes become activated. This is characterized by changes in the gene expression profiles and high levels of GFAP [57]. In addition to this, exposure to corticosterone for short (6-24 h) and prolonged (3 weeks) period in astrocytic cultures show an increase in GFAP mRNA levels and a decrease when astrocytes were co-cultured with neurons [57]. This supports the findings that GCs may also regulate the synthesis of GFAP. Further, it also suggests the crosstalk between neuron and astrocytes to withstand deleterious effects of corticosterone. Initial in-vivo studies on rats have shown decreased hippocampal and cortical GFAP mRNA levels [58] while immunohistochemical studies have shown increased GFAP immunoreactivity in a dose and brain-region specific manner following chronic corticosterone treatment [59]. The complex nature of corticosteroids regulating GFAP during stress can be understood wherein 6 days of stressful activity leads to 30% increase in hippocampal GFAP-immunoreactive astrocytes [60] and at the same instance, the studies on adult rats exposed to early-life stress showed reduced density of GFAP-immunoreactive astrocytes in various limbic regions of brain [61]. This GFAP binary response may suggest a primary astrocyte-mediated neural protection, which may subsequently turn neurotoxic depending on the dose, which will affect the brain region as well as the time of exposure to stress.
Astrocytes express most of the receptors and additionally, ion channels found in neurons are involved in recycling and eliminating glutamate from synapses, thereby contributing to glutamatergic synaptic transmission [62]. The glial transporters help glutamate uptake and conversion to glutamine with the help of enzyme glutamine synthase (GS), which is responsive towards GCs [63]. A current hypothesis states that excessive extra synaptic glutamate leads to the atrophy of apical dendrites seen in hippocampal pyramidal neurons in stressed rats [64]. Studies aiming to understand the regulation of GS have shown glucocorticoid-mediated regulations of glial GS during stress [63]. Likewise, chronic stress showed up-regulation of the hippocampal glial glutamate transporter (GLT-1) (Figure 1F) [65]. This suggests that glutamate cycling is regulated by corticosteroids by induction of GS and GLT-1 expression in a time-dependent manner in distinct subpopulation of astrocytes. The hypothesis that GCs inhibit glucose uptake was substantially evidenced by a study in which it was shown that chronic mild stress exacerbates the consequences of chronic cerebral hypoperfusion [66]. This results in a lack of energy to neurons and astrocytes for high-affinity glutamate reuptake and thus an increase in vulnerability of the brain [67].
Astrocytes lie in proximity of BBB and play an important role in its maintenance by interacting with endothelial cells. On this regard, it would be important to emphasize that modulation of the barrier “tightness” is a result of complex interaction between GCs and cells of neurovascular units including astrocytes [68].
As stated earlier, S100β found primarily in the cytoplasm of astrocytes, and is involved in glia-neuron signaling [69]. The S100β regulates a variety of intra and extracellular functions such as cell growth, metabolism, calcium homeostasis and synaptic plasticity [69]. This suggests the possible role of astrocytes in regulating neuronal synaptic plasticity. Furthermore, reports suggest that S100β concentration was reduced after maternal administration of betamethasone (a synthetic glucocorticoid) in the hippocampus and serum of the neonate rat [70]. This suggests that increased GCs or chronic stress may reduce the expression or function of S100β. In an another study, stress exposure increased S100β concentration in cerebrospinal fluid (CSF) after acute predator stress [71] and chronic restraint stress [72]. These findings reveal the biphasic response of S100β to stress. This observation was also supported by another study where the astrocyte cultures were exposed to dexamethasone (Table 1)[73].
| Name of the marker | Function | References |
| Glial Fibrillary Acidic Protein (GFAP) | Cytoskeletal protein | [58] |
| S100β | astroglia-specific neurotrophic factor | [82,83] |
| Nerve growth factor (NGF) | Neurotrophic factor | [73] |
| Basic fibroblast growth factor (bFGF) | Neurotrophic factor | [73] |
| N-myc downstream- regulated Gene (Ndrg2) | Cell differentiation factor | [84] |
| Glutamine synthetase (GS) | Recycling of the glutamate | [37,53,63] |
| Glial glutamate transporter (GLT-1) | Recycling of the glutamate | [37,53,63] |
 DownLoad: CSV
DownLoad: CSVAstrocytes may coordinate their function through Ca2+ excitability and subsequent signaling that have also been implicated in depressive disorders [74]. Surprisingly, astrocytic calcium signaling is regulated by GCs [75]. This apparently suggests the role of GCs in the modulation of glial calcium cell signaling during depression and anxiety. Additionally, the GCs are critical regulators of brain development and brain aging. Rat astrocyte primary culture study showed that dexamethasone increases intracellular and membrane-associated lipocortin-1 (annexin-1) [76] while it increased and reduced expression of nerve growth factor (NGF) in cultured neurons and astrocytes respectively [77].
Recent work focused on how the astrocytes intervene in the GC-induced stress and its probable mechanism. It has been shown that 5' AMP-activated protein kinase (AMPK) may mediate down regulation of GRs in astrocytes of rat PFC [78]. Also, GRs in astrocytes, as a critical stress-responding transcriptional factor, may mediate stress-induced adaptation via regulating the expression of astrocyte-derived neurotrophic factors. Researchers also found that the chronic mild stress-induced decrease in brain-derived neurotrophic factor (BDNF) and Cx43 causing astrocytic dysfunction by forming abnormality in gap junctions in the PFC of rats [79,80]. Some recent studies in emotional processing show astrocytes' crucial role, along with neurons, in the hippocampus, PFC, and amygdala [81]. Thus current astrobiology research suggests an evolving role of neuro-astroglial communication in the modulation of depression, opening new avenues of therapies against depressive disorders.
The studies over recent years have started unveiling the role of astrocytes in stress and their response to GCs release thus shifting our understanding from neuron-centric theories towards the role of these glial cells in the neurobiology of stress. However, the role of released GCs in response to stress and its effects on astrocytes need to be explored further in for a better understanding of stress- or glucocorticoid- related brain disorders.
The authors sincerely thank Mr. Michael Klug, Department of Mathematics, UC Berkeley, USA, for his diligent proof-reading of this manuscript.
The authors declare no conflict of interests.
| [1] | Volterra A, Magistretti PJ, Haydon PG (2002) The tripartite synapse: Glia in synaptic transmission. New York: Oxford University Press. |
| [2] |
Pelvig DP, Pakkenberg H, Stark AK, et al. (2008) Neocortical glial cell numbers in human brains. Neurobiol Aging 29: 1754–1762. doi: 10.1016/j.neurobiolaging.2007.04.013

|
| [3] |
Ge WP, Jia JM (2016) Local production of astrocytes in the cerebral cortex. Neuroscience 323: 3–9. doi: 10.1016/j.neuroscience.2015.08.057
|
| [4] |
Hertz L (2008) Bioenergetics of cerebral ischemia: A cellular perspective. Neuropharmacology 55: 289–309. doi: 10.1016/j.neuropharm.2008.05.023
|
| [5] |
Bröer S, Brookes N (2001) Transfer of glutamine between astrocytes and neurons. J Neurochem 77: 705–719. doi: 10.1046/j.1471-4159.2001.00322.x
|
| [6] |
Halassa MM, Fellin T, Haydon PG (2007) The tripartite synapse: Roles for gliotransmission in health and disease. Trends Mol Med 13: 54–63. doi: 10.1016/j.molmed.2006.12.005
|
| [7] |
Nualart-Marti A, Solsona C, Fields RD (2013) Gap junction communication in myelinating glia. Biochim Biophys Acta 1828: 69–78. doi: 10.1016/j.bbamem.2012.01.024
|
| [8] | Cabezas R, Avila M, Gonzalez J, et al. (2014) Astrocytic modulation of blood brain barrier: Perspectives on Parkinson's disease. Front Cell Neurosci 8: 211. |
| [9] | Sapolsky RM, Romero LM, Munck AU (2000) How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev 21: 55–89. |
| [10] |
Chrousos GP (2009) Stress and disorders of the stress system. Nat Rev Endocrinol 5: 374–381. doi: 10.1038/nrendo.2009.106
|
| [11] |
De Kloet ER, Van Acker SA, Sibug RM, et al. (2000) Brain mineralocorticoid receptors and centrally regulated functions. Kidney Int 57: 1329–1336. doi: 10.1046/j.1523-1755.2000.00971.x
|
| [12] |
Carter BS, Meng F, Thompson RC (2012) Glucocorticoid treatment of astrocytes results in temporally dynamic transcriptome regulation and astrocyte-enriched mRNA changes in vitro. Physiol Genomics 44: 1188–1200. doi: 10.1152/physiolgenomics.00097.2012
|
| [13] |
Mcewen BS (2005) Glucocorticoids, depression, and mood disorders: Structural remodeling in the brain. Metabolism 54: 20–23. doi: 10.1016/j.metabol.2005.01.008
|
| [14] |
Green KN, Billings LM, Roozendaal B, et al. (2006) Glucocorticoids increase amyloid-beta and tau pathology in a mouse model of Alzheimer's disease. J Neurosci 26: 9047–9056. doi: 10.1523/JNEUROSCI.2797-06.2006
|
| [15] |
Newell-Price J, Bertagna X, Grossman AB, et al. (2006) Cushing's syndrome. Lancet 367: 1605–1617. doi: 10.1016/S0140-6736(06)68699-6
|
| [16] |
Cobb J, O'Neill K, Milner J, et al. (2016) Density of GFAP-immunoreactive astrocytes is decreased in left hippocampi in major depressive disorder. Neuroscience 316: 209–220. doi: 10.1016/j.neuroscience.2015.12.044
|
| [17] | Vyas S, Rodrigues AJ, Silva JM, et al. (2016) Chronic stress and glucocorticoids: From neuronal plasticity to neurodegeneration. Neural Plast 2016: 6391686. |
| [18] |
Volterra A, Meldolesi J (2005) Astrocytes, from brain glue to communication elements: The revolution continues. Nat Rev Neurosci 6: 626–640. doi: 10.1038/nrn1722
|
| [19] | Koyama Y (2015) Functional alterations of astrocytes in mental disorders: Pharmacological significance as a drug target. Front Cell Neurosci 9: 261. |
| [20] | Carter BS (2013) Glucocorticoid regulation of the astrocyte transcriptome in vitro and in vivo. University of Michigan. |
| [21] |
Conti F, Debiasi S, Minelli A, et al. (1996) Expression of NR1 and NR2A/B subunits of the NMDA receptor in cortical astrocytes. Glia 17: 254–258. doi: 10.1002/(SICI)1098-1136(199607)17:3<254::AID-GLIA7>3.0.CO;2-0
|
| [22] |
Rajkowska G, Miguel-Hidalgo JJ, Wei J, et al. (1999) Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression. Biol Psychiatry 45: 1085–1098. doi: 10.1016/S0006-3223(99)00041-4
|
| [23] |
Stockmeier CA, Mahajan GJ, Konick LC, et al. (2004) Cellular changes in the postmortem hippocampus in major depression. Biol Psychiatry 56: 640–650. doi: 10.1016/j.biopsych.2004.08.022
|
| [24] |
Rajkowska G, Stockmeier CA (2013) Astrocyte pathology in major depressive disorder: Insights from human postmortem brain tissue. Curr Drug Targets 14: 1225–1236. doi: 10.2174/13894501113149990156
|
| [25] |
Verkhratsky A, Rodriguez JJ, Parpura V (2013) Astroglia in neurological diseases. Future Neurol 8: 149–158. doi: 10.2217/fnl.12.90
|
| [26] |
Banasr M, Duman RS (2008) Glial Loss in the prefrontal cortex is sufficient to induce depressive-like behaviors. Biol Psychiatry 64: 863–870. doi: 10.1016/j.biopsych.2008.06.008
|
| [27] | Czeh B, Simon M, Schmelting B, et al. (2005) Astroglial plasticity in the hippocampus is affected by chronic psychosocial stress and concomitant fluoxetine treatment. Neuropsychopharmacology 31: 1616–1626. |
| [28] |
Banasr M, Chowdhury GMI, Terwilliger R, et al. (2010) Glial pathology in an animal model of depression: Reversal of stress-induced cellular, metabolic and behavioral deficits by the glutamate-modulating drug riluzole. Mol Psychiatry 15: 501–511. doi: 10.1038/mp.2008.106
|
| [29] |
Messing A, Brenner M (2003) GFAP: Functional implications gleaned from studies of genetically engineered mice. Glia 43: 87–90. doi: 10.1002/glia.10219
|
| [30] |
Miguel-Hidalgo JJ, Baucom C, Dilley G, et al. (2000) Glial fibrillary acidic protein immunoreactivity in the prefrontal cortex distinguishes younger from older adults in major depressive disorder. Biol Psychiatry 48: 861–873. doi: 10.1016/S0006-3223(00)00999-9
|
| [31] |
Bowley MP, Drevets WC, Ongur D, et al. (2002) Low glial numbers in the amygdala in major depressive disorder. Biol Psychiatry 52: 404–412. doi: 10.1016/S0006-3223(02)01404-X
|
| [32] | Polyakova M, Sander C, Arelin K, et al. (2015) First evidence for glial pathology in late life minor depression: S100B is increased in males with minor depression. Front Cell Neurosci 9: 406. |
| [33] |
Benton CS, Miller BH, Skwerer S, et al. (2012) Evaluating genetic markers and neurobiochemical analytes for fluoxetine response using a panel of mouse inbred strains. Psychopharmacology 221: 297–315. doi: 10.1007/s00213-011-2574-z
|
| [34] |
Chandley MJ, Szebeni K, Szebeni A, et al. (2013) Gene expression deficits in pontine locus coeruleus astrocytes in men with major depressive disorder. J Psychiatry Neurosci 38: 276–284. doi: 10.1503/jpn.120110
|
| [35] |
Bernard R, Kerman IA, Thompson RC, et al. (2011) Altered expression of glutamate signaling, growth factor, and glia genes in the locus coeruleus of patients with major depression. Mol Psychiatry 16: 634–646. doi: 10.1038/mp.2010.44
|
| [36] |
Seifert G, Schilling K, Steinhäuser C (2006) Astrocyte dysfunction in neurological disorders: A molecular perspective. Nat Rev Neurosci 7: 194. doi: 10.1038/nrn1870
|
| [37] |
Choudary P, Molnar M, Evans S, et al. (2005) Altered cortical glutamatergic and GABAergic signal transmission with glial involvement in depression. Proc Natl Acad Sci 102: 15653–15658. doi: 10.1073/pnas.0507901102
|
| [38] |
Sun JD, Liu Y, Yuan YH, et al. (2012) Gap junction dysfunction in the prefrontal cortex induces depressive-like behaviors in rats. Neuropsychopharmacology 37: 1305–1320. doi: 10.1038/npp.2011.319
|
| [39] |
Bechtholt-Gompf AJ, Walther HV, Adams MA, et al. (2010) Blockade of astrocytic glutamate uptake in rats induces signs of anhedonia and impaired spatial memory. Neuropsychopharmacology 35: 2049–2059. doi: 10.1038/npp.2010.74
|
| [40] |
de Vasconcellos-Bittencourt AP, Vendite DA, Nassif M, et al. (2011) Chronic stress and lithium treatments alter hippocampal glutamate uptake and release in the rat and potentiate necrotic cellular death after oxygen and glucose deprivation. Neurochem Res 36: 793–800. doi: 10.1007/s11064-011-0404-7
|
| [41] |
Almeida RF, Thomazi AP, Godinho GF, et al. (2010) Effects of depressive-like behavior of rats on brain glutamate uptake. Neurochem Res 35: 1164–1171. doi: 10.1007/s11064-010-0169-4
|
| [42] |
Bechtholt-Gompf AJ, Walther HV, Adams MA, et al. (2010) Blockade of astrocytic glutamate uptake in rats induces signs of anhedonia and impaired spatial memory. Neuropsychopharmacology 35: 2049–2059. doi: 10.1038/npp.2010.74
|
| [43] | Lapidus KA, Soleimani L, Murrough JW (2013) Novel glutamatergic drugs for the treatment of mood disorders. Neuropsychiatr Dis Treat 9: 1101–1112. |
| [44] |
Sanacora G, Treccani G, Popoli M (2012) Towards a glutamate hypothesis of depression: An emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacology 62: 63–77. doi: 10.1016/j.neuropharm.2011.07.036
|
| [45] |
Paslakis G, Gass P, Deuschle M (2011) [The role of the glutamatergic system in pathophysiology and pharmacotherapy for depression: Preclinical and clinical data]. Fortschr Neurol Psychiatr 79: 204–212. doi: 10.1055/s-0029-1245770
|
| [46] | Rial D, Lemos C, Pinheiro H, et al. (2015) Depression as a glial-based synaptic dysfunction. Front Cell Neurosci 9: 521. |
| [47] | Jo WK, Zhang Y, Emrich HM, et al. (2015) Glia in the cytokine-mediated onset of depression: fine tuning the immune response. Front Cell Neurosci 9: 268. |
| [48] | Brites D, Fernandes A (2015) Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front Cell Neurosci 9: 476. |
| [49] | Quesseveur G, Portal B, Basile JA, et al. (2015) Attenuated Levels of Hippocampal Connexin 43 and its Phosphorylation Correlate with Antidepressant- and Anxiolytic-Like Activities in Mice. Front Cell Neurosci 9: 490. |
| [50] | Jeanson T, Pondaven A, Ezan P, et al. (2015) Antidepressants Impact Connexin 43 Channel Functions in Astrocytes. Front Cell Neurosci 9: 495. |
| [51] |
Fuchs E, Flugge G (2003) Chronic social stress: Effects on limbic brain structures. Physiol Behav 79: 417–427. doi: 10.1016/S0031-9384(03)00161-6
|
| [52] |
Gould E, Tanapat P (1999) Stress and hippocampal neurogenesis. Biol Psychiatry 46: 1472–1479. doi: 10.1016/S0006-3223(99)00247-4
|
| [53] |
Rajkowska G, Miguel-Hidalgo JJ (2007) Gliogenesis and glial pathology in depression. CNS Neurol Disord Drug Targets 6: 219–233. doi: 10.2174/187152707780619326
|
| [54] |
Wang Q, Verweij EW, Krugers HJ, et al. (2014) Distribution of the glucocorticoid receptor in the human amygdala; changes in mood disorder patients. Brain Struct Funct 219: 1615–1626. doi: 10.1007/s00429-013-0589-4
|
| [55] |
Czeh B, Muller-Keuker JI, Rygula R, et al. (2007) Chronic social stress inhibits cell proliferation in the adult medial prefrontal cortex: Hemispheric asymmetry and reversal by fluoxetine treatment. Neuropsychopharmacology 32: 1490–1503. doi: 10.1038/sj.npp.1301275
|
| [56] |
Pekny M, Nilsson M (2005) Astrocyte activation and reactive gliosis. Glia 50: 427–434. doi: 10.1002/glia.20207
|
| [57] |
Rozovsky I, Laping NJ, Krohn K, et al. (1995) Transcriptional regulation of glial fibrillary acidic protein by corticosterone in rat astrocytes in vitro is influenced by the duration of time in culture and by astrocyte-neuron interactions. Endocrinology 136: 2066–2073. doi: 10.1210/endo.136.5.7720656
|
| [58] |
O'Callaghan JP, Brinton RE, Mcewen BS (1991) Glucocorticoids regulate the synthesis of glial fibrillary acidic protein in intact and adrenalectomized rats but do not affect its expression following brain injury. J Neurochem 57: 860–869. doi: 10.1111/j.1471-4159.1991.tb08230.x
|
| [59] | Bridges N, Slais K, Sykova E (2008) The effects of chronic corticosterone on hippocampal astrocyte numbers: A comparison of male and female Wistar rats. Acta Neurobiol Exp 68: 131–138. |
| [60] |
Lambert KG, Gerecke KM, Quadros PS, et al. (2000) Activity-stress increases density of GFAP-immunoreactive astrocytes in the rat hippocampus. Stress 3: 275–284. doi: 10.3109/10253890009001133
|
| [61] |
Leventopoulos M, Ruedi-Bettschen D, Knuesel I, et al. (2007) Long-term effects of early life deprivation on brain glia in Fischer rats. Brain Res 1142: 119–126. doi: 10.1016/j.brainres.2007.01.039
|
| [62] |
Fields RD, Stevens-Graham B (2002) New insights into neuron-glia communication. Science 298: 556–562. doi: 10.1126/science.298.5593.556
|
| [63] |
Vardimon L, Ben-Dror I, Avisar N, et al. (1999) Glucocorticoid control of glial gene expression. J Neurobiol 40: 513–527. doi: 10.1002/(SICI)1097-4695(19990915)40:4<513::AID-NEU8>3.0.CO;2-D
|
| [64] |
Conrad CD (2006) What is the functional significance of chronic stress-induced CA3 dendritic retraction within the hippocampus? Behav Cogn Neurosci Rev 5: 41–60. doi: 10.1177/1534582306289043
|
| [65] |
Autry AE, Grillo CA, Piroli GG, et al. (2006) Glucocorticoid regulation of GLT-1 glutamate transporter isoform expression in the rat hippocampus. Neuroendocrinology 83: 371–379. doi: 10.1159/000096092
|
| [66] |
Horner HC, Packan DR, Sapolsky RM (1990) Glucocorticoids inhibit glucose transport in cultured hippocampal neurons and glia. Neuroendocrinology 52: 57–64. doi: 10.1159/000125539
|
| [67] |
Ritchie LJ, De Butte M, Pappas BA (2004) Chronic mild stress exacerbates the effects of permanent bilateral common carotid artery occlusion on CA1 neurons. Brain Res 1014: 228–235. doi: 10.1016/j.brainres.2004.04.036
|
| [68] |
Kroll S, El-Gindi J, Thanabalasundaram G, et al. (2009) Control of the blood-brain barrier by glucocorticoids and the cells of the neurovascular unit. Ann N Y Acad Sci 1165: 228–239. doi: 10.1111/j.1749-6632.2009.04040.x
|
| [69] |
Donato R (2001) S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int J Biochem Cell Biol 33: 637–668. doi: 10.1016/S1357-2725(01)00046-2
|
| [70] |
Bruschettini M, van den Hove DL, Gazzolo D, et al. (2005) A single course of antenatal betamethasone reduces neurotrophic factor S100B concentration in the hippocampus and serum in the neonatal rat. Brain Res Dev Brain Res 159: 113–118. doi: 10.1016/j.devbrainres.2005.07.003
|
| [71] |
Margis R, Zanatto VC, Tramontina F, et al. (2004) Changes in S100B cerebrospinal fluid levels of rats subjected to predator stress. Brain Res 1028: 213–218. doi: 10.1016/j.brainres.2004.09.013
|
| [72] |
Scaccianoce S, Del BP, Pannitteri G, et al. (2004) Relationship between stress and circulating levels of S100B protein. Brain Res 1004: 208–211. doi: 10.1016/j.brainres.2004.01.028
|
| [73] |
Niu H, Hinkle DA, Wise PM (1997) Dexamethasone regulates basic fibroblast growth factor, nerve growth factor and S100beta expression in cultured hippocampal astrocytes. Brain Res Mol Brain Res 51: 97–105. doi: 10.1016/S0169-328X(97)00221-0
|
| [74] |
Bazargani N, Attwell D (2016) Astrocyte calcium signaling: the third wave. Nat Neurosci 19: 182–189. doi: 10.1038/nn.4201
|
| [75] |
Simard M, Couldwell WT, Zhang W, et al. (1999) Glucocorticoids-potent modulators of astrocytic calcium signaling. Glia 28: 1–12. doi: 10.1002/(SICI)1098-1136(199910)28:1<1::AID-GLIA1>3.0.CO;2-4
|
| [76] |
Mcleod JD, Bolton C (1995) Dexamethasone induces an increase in intracellular and membrane-associated lipocortin-1 (annexin-1) in rat astrocyte primary cultures. Cell Mol Neurobiol 15: 193–205. doi: 10.1007/BF02073328
|
| [77] |
Lindholm D, Castren E, Hengerer B, et al. (1992) Differential Regulation of Nerve Growth Factor (NGF) Synthesis in Neurons and Astrocytes by Glucocorticoid Hormones. Eur J Neurosci 4: 404–410. doi: 10.1111/j.1460-9568.1992.tb00889.x
|
| [78] |
Yuan SY, Liu J, Zhou J, et al. (2016) AMPK Mediates Glucocorticoids Stress-Induced Downregulation of the Glucocorticoid Receptor in Cultured Rat Prefrontal Cortical Astrocytes. PLoS One 11: e0159513. doi: 10.1371/journal.pone.0159513
|
| [79] |
Chen J, Wang ZZ, Zuo W, et al. (2016) Effects of chronic mild stress on behavioral and neurobiological parameters-Role of glucocorticoid. Horm Behav 78: 150–159. doi: 10.1016/j.yhbeh.2015.11.006
|
| [80] |
Karisetty BC, Joshi PC, Kumar A, et al. (2017) Sex differences in the effect of chronic mild stress on mouse prefrontal cortical BDNF levels: A role of major ovarian hormones. Neuroscience 356: 89–101. doi: 10.1016/j.neuroscience.2017.05.020
|
| [81] |
Bender CL, Calfa GD, Molina VA (2016) Astrocyte plasticity induced by emotional stress: A new partner in psychiatric physiopathology? Prog Neuropsychopharmacol Biol Psychiatry 65: 68–77. doi: 10.1016/j.pnpbp.2015.08.005
|
| [82] |
Anacker C, Cattaneo A, Luoni A, et al. (2013) Glucocorticoid-related molecular signaling pathways regulating hippocampal neurogenesis. Neuropsychopharmacology 38: 872–883. doi: 10.1038/npp.2012.253
|
| [83] |
Van DL, Steinbusch HW, Bruschettini M, et al. (2006) Prenatal stress reduces S100B in the neonatal rat hippocampus. Neuroreport 17: 1077–1080. doi: 10.1097/01.wnr.0000223391.74575.c9
|
| [84] |
Nichols NR, Agolley D, Zieba M, et al. (2005) Glucocorticoid regulation of glial responses during hippocampal neurodegeneration and regeneration. Brain Res Rev 48: 287–301. doi: 10.1016/j.brainresrev.2004.12.019
|
| 1. | Zuleide M. Ignácio, Renato S. da Silva, Marcos E. Plissari, João Quevedo, Gislaine Z. Réus, Physical Exercise and Neuroinflammation in Major Depressive Disorder, 2019, 56, 0893-7648, 8323, 10.1007/s12035-019-01670-1 | |
| 2. | Yi-Hung Chen, Sheng-Yun Xie, Chao-Wei Chen, Dah-Yuu Lu, Electroacupuncture improves repeated social defeat stress-elicited social avoidance and anxiety-like behaviors by reducing Lipocalin-2 in the hippocampus, 2021, 14, 1756-6606, 10.1186/s13041-021-00860-0 | |
| 3. | Εvgenia Dandi, Paschalis Theotokis, Maria Christina Petri, Vaia Sideropoulou, Evangelia Spandou, Despina A. Tata, Environmental enrichment initiated in adolescence restores the reduced expression of synaptophysin and GFAP in the hippocampus of chronically stressed rats in a sex‐specific manner, 2023, 65, 0012-1630, 10.1002/dev.22422 | |
| 4. | José Javier Miguel-Hidalgo, Neuroprotective astroglial response to neural damage and its relevance to affective disorders, 2023, 328, 10.37349/ent.2023.00054 | |
| 5. | Ana Laura Reyes-Ábalos, Magdalena Álvarez-Zabaleta, Silvia Olivera-Bravo, María Vittoria Di Tomaso, Tejeshwar Rao, Acute Genetic Damage Induced by Ethanol and Corticosterone Seems to Modulate Hippocampal Astrocyte Signaling, 2024, 2024, 1687-8884, 1, 10.1155/2024/5524487 | |
| 6. | Jingqi Le, Wangyan Deng, Tao Le, Music Therapy in Depression: Exploring Mechanisms and Efficacy in Rat Models, 2025, 15, 2076-3425, 338, 10.3390/brainsci15040338 |

Figures(1) / Tables(1)

Pranav Chintamani Joshi, Sugato Benerjee. Effects of glucocorticoids in depression: Role of astrocytes[J]. AIMS Neuroscience, 2018, 5(3): 200-210. doi: 10.3934/Neuroscience.2018.3.200

| Name of the marker | Function | References |
| Glial Fibrillary Acidic Protein (GFAP) | Cytoskeletal protein | [58] |
| S100β | astroglia-specific neurotrophic factor | [82,83] |
| Nerve growth factor (NGF) | Neurotrophic factor | [73] |
| Basic fibroblast growth factor (bFGF) | Neurotrophic factor | [73] |
| N-myc downstream- regulated Gene (Ndrg2) | Cell differentiation factor | [84] |
| Glutamine synthetase (GS) | Recycling of the glutamate | [37,53,63] |
| Glial glutamate transporter (GLT-1) | Recycling of the glutamate | [37,53,63] |
DownLoad: CSV| Name of the marker | Function | References |
| Glial Fibrillary Acidic Protein (GFAP) | Cytoskeletal protein | [58] |
| S100β | astroglia-specific neurotrophic factor | [82,83] |
| Nerve growth factor (NGF) | Neurotrophic factor | [73] |
| Basic fibroblast growth factor (bFGF) | Neurotrophic factor | [73] |
| N-myc downstream- regulated Gene (Ndrg2) | Cell differentiation factor | [84] |
| Glutamine synthetase (GS) | Recycling of the glutamate | [37,53,63] |
| Glial glutamate transporter (GLT-1) | Recycling of the glutamate | [37,53,63] |





