Citation: Sabrine Louhaichi, Mariem Salhi, Anissa Berraïes, Besma Hamdi, Jamel Ammar, Kamel Hamzaoui, Agnès Hamzaoui. Co-inhibitory receptors in female asthmatic patients: Correlation with IL-17 and IL-26[J]. AIMS Allergy and Immunology, 2018, 2(1): 10-23. doi: 10.3934/Allergy.2018.1.10

| [1] |
Cosmi L, Liotta F, Maggi L, et al. (2017) Role of Type 2 innate lymphoid cellsin allergic diseases. Curr Allergy Asthm R 17: 66–72. doi: 10.1007/s11882-017-0735-9

|
| [2] |
Galowitz S, Chang C (2015) Immunobiology of critical pediatric asthma. Clin Rev Allerg Immu 48: 84–96. doi: 10.1007/s12016-014-8409-z
|
| [3] |
Charrad R, Berraïes A, Hamdi B, et al. (2016) Anti-inflammatory activity of IL-37 in asthmatic children: Correlation with inflammatory cytokines TNF-α, IL-β, IL-6 and IL-17A. Immunobiology 221: 182–187. doi: 10.1016/j.imbio.2015.09.009
|
| [4] |
Bernard A, Lamy AL, Alberti I (2002) The two-signal model of T-cell activation after 30 years. Transplantation 73: S31–S35. doi: 10.1097/00007890-200201151-00011
|
| [5] | Boulougouris G, Mcleod JD, Patel YI, et al. (1998) Positive and negative regulation of human T cell activation mediated by the CTLA-4/CD28 ligand CD80. J Immunol 161: 3919–3924. |
| [6] |
Freeman GJ, Long AJ, Iwai Y, et al. (2000) Engagement of the PD-1immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med 192: 1027–1034. doi: 10.1084/jem.192.7.1027
|
| [7] | Wang XF, Chen YJ, Wang Q, et al. (2007) Distinct expression and inhibitory function of B and T lymphocyte attenuator on human T cells. HLA 69: 145–153. |
| [8] |
Waterhouse P, Penninger JM, Timms E, et al. (1995) Lymphoproliferative disorders with early lethality in mice deficient in Ctla-4. Science 270: 985–988. doi: 10.1126/science.270.5238.985
|
| [9] |
Boomer JS, Jennifer SS, Hotchkiss RS, et al. (2012) A prospective analysis of lymphocyte phenotypeand function over the course of acute sepsis. Crit Care 16: R112. doi: 10.1186/cc11404
|
| [10] |
Van WC, Bateman ED, Bousquet J, et al. (2008) Asthma management pocket reference 2008. Allergy 63: 997–1004. doi: 10.1111/j.1398-9995.2008.01643.x
|
| [11] |
Berraïes A, Hamdi B, Ammar J, et al. (2016) Increased Expression of Thymic Stromal Lymphopoietin in induced sputum from asthmatic children. Immunol Lett 178: 85–91. doi: 10.1016/j.imlet.2016.08.004
|
| [12] |
Moermans C, Heinen V, Nguyen M, et al. (2011) Local and systemic cellular inflammation and cytokine release in chronic obstructive pulmonary disease. Cytokine 56: 298–304. doi: 10.1016/j.cyto.2011.07.010
|
| [13] |
Delvaux M, Henket M, Lau L, et al. (2004) Nebulised salbutamol administered during sputum induction improves bronchoprotection in patients with asthma. Thorax 59: 111–115. doi: 10.1136/thorax.2003.011130
|
| [14] |
Kaabachi W, Berraïes A, Dhifallh IB, et al. (2017) Interleukin-26 is overexpressed in Behçet's disease and enhances Th17 related -cytokines. Immunol Lett 190: 177–184. doi: 10.1016/j.imlet.2017.08.008
|
| [15] |
Wang X, Zhu G, Huang Z, et al. (2012) Application of monoclonal antibodies in a sandwich enzyme-linked immunosorbent assay for identification and detection of soluble human B and T lymphocyte attenuator. Hybridoma 31: 417–423. doi: 10.1089/hyb.2012.0052
|
| [16] |
Cai G, Freeman GJ (2009) The CD160, BTLA, LIGHT/HVEM pathway: a bidirectional switch regulating T-cell activation. Immunol Rev 229: 244–258. doi: 10.1111/j.1600-065X.2009.00783.x
|
| [17] |
Oaks MK, Hallett KM, Penwell RT, et al. (2000) A native soluble form of CTLA-4. Cell Immunol 201: 144–153. doi: 10.1006/cimm.2000.1649
|
| [18] |
Nielsen C, Ohm-Laursen L, Barington T, et al. (2005) Alternative splice variants of the human PD-1 gene. Cell Immunol 235: 109–116. doi: 10.1016/j.cellimm.2005.07.007
|
| [19] |
Magistrelli G, Jeannin P, Herbault N, et al. (1999) A soluble form of CTLA-4 generated by alternative splicing is expressed by nonstimulated human T cells. Eur J Immunol 29: 3596–3602. doi: 10.1002/(SICI)1521-4141(199911)29:11<3596::AID-IMMU3596>3.0.CO;2-Y
|
| [20] | Chesné J, Braza F, Mahay G, et al. (2014) IL-17 in severe asthma. Where do we stand? Am J Respir Crit Care Med 190: 1094–1101. |
| [21] |
Bullens DM, Truyen E, Coteur L, et al. (2006) IL-17 mRNA in sputum of asthmatic patients: linking T cell driven inflammation and granulocytic influx? Resp Res 7: 135. doi: 10.1186/1465-9921-7-135
|
| [22] |
Wu H, Miao M, Zhang G, et al. (2009) Soluble PD-1 isassociated with aberrant regulation of T cells activation in aplasticanemia. Immunol Invest 38: 408–421. doi: 10.1080/08820130902912332
|
| [23] |
Song MY, Park SH, Nam HJ, et al. (2011) Enhancementof vaccine-induced primary and memory CD8(+) T-cell responsesby soluble PD-1. J Immunother 34: 297–306. doi: 10.1097/CJI.0b013e318210ed0e
|
| [24] |
Kuipers H, Muskens F, Willart M, et al. (2006) Contribution of the PD-1 ligands/PD-1 signaling pathway to dendritic cell-mediated CD4+ T cell activation. Eur J Immunol 36: 2472–2482. doi: 10.1002/eji.200635978
|
| [25] | Hasegawa T, Uga H, Mori A, et al. (2017) Increased serum IL-17A and Th2 cytokinelevels in patients with severe uncontrolled asthma. Eur Cytokine Netw 28: 8–18. |
| [26] |
Pyle CJ, Uwadiae FI, Swieboda DP, et al. (2017) Early IL-6 signalling promotesIL-27 dependent maturation of regulatory T cells in the lungs and resolution ofviral immunopathology. PLoS Pathog 13: e1006640. doi: 10.1371/journal.ppat.1006640
|
| [27] |
Maalmi H, Beraies A, Charad R, et al. (2014) IL-17A and IL-17F genes variants and susceptibility to childhood asthma in Tunisia. J Asthma 51: 348–354. doi: 10.3109/02770903.2013.876647
|
| [28] | Charrad R, Kaabachi W, Berraies A, et al. (2017) IL-33 gene variants and protein expression in pediatric Tunisian asthmatic patients. Cytokine 11: 32. |
| [29] |
Chen JH, Qin L, Shi YY, et al. (2016) IL-17 protein levels in both induced sputum and plasma are increased in stable but not acute asthma individuals with obesity. Resp Med 121: 48–58. doi: 10.1016/j.rmed.2016.10.018
|
| [30] |
Lindén A, Dahlén B (2014) Interleukin-17 cytokine signalling in patients with asthma. Eur Respir J 44: 1319–1331. doi: 10.1183/09031936.00002314
|
| [31] |
Ricciardolo FL, Sorbello V, Folino A, et al. (2017) Identification of IL-17F/frequent exacerbator endotype in asthma. J Allergy Clin Immunol 140: 395–406. doi: 10.1016/j.jaci.2016.10.034
|
| [32] |
Griffiths KL, Khader SA (2014) Bringing in the cavalry: IL-26 mediates neutrophil recruitment to the lungs. Am J Respir Crit Care Med 190: 1079–1080. doi: 10.1164/rccm.201410-1870ED
|
| [33] |
Ohnuma K, Hatano R, Aune TM, et al. (2015). Regulation of pulmonary graft-versus-host disease by IL- 26+CD26+CD4 T lymphocytes. J Immunol 194: 3697–3712. doi: 10.4049/jimmunol.1402785
|
| [34] |
Burlingham WJ, Love RB, Jankowskagan E, et al. (2007) IL-17-dependent cellular immunity to collagen Type V predisposes to obliterative bronchiolitis in human lung transplants. J Clin Invest 117: 3498–3506. doi: 10.1172/JCI28031
|
| [35] |
Che KF, Tengvall S, Levanen B, et al. (2014) Interleukin-26 in antibacterial host defense of human lungs. Effects on neutrophil mobilization. Am J Respir Crit Care Med 190: 1022–1031. doi: 10.1164/rccm.201404-0689OC
|
| [36] |
Elgaili AA, Nzungize L, Duan X, et al. (2016) Interleukin-10 family and tuberculosis: Old story renewed. Int J Biol Sci 12: 710–717. doi: 10.7150/ijbs.13881
|
| [37] |
Charrad R, Kaabachi W, Rafrafi A, et al. (2017) IL-8 gene variants and expression in childhood asthma. Lung 195: 1–9. doi: 10.1007/s00408-016-9960-6
|
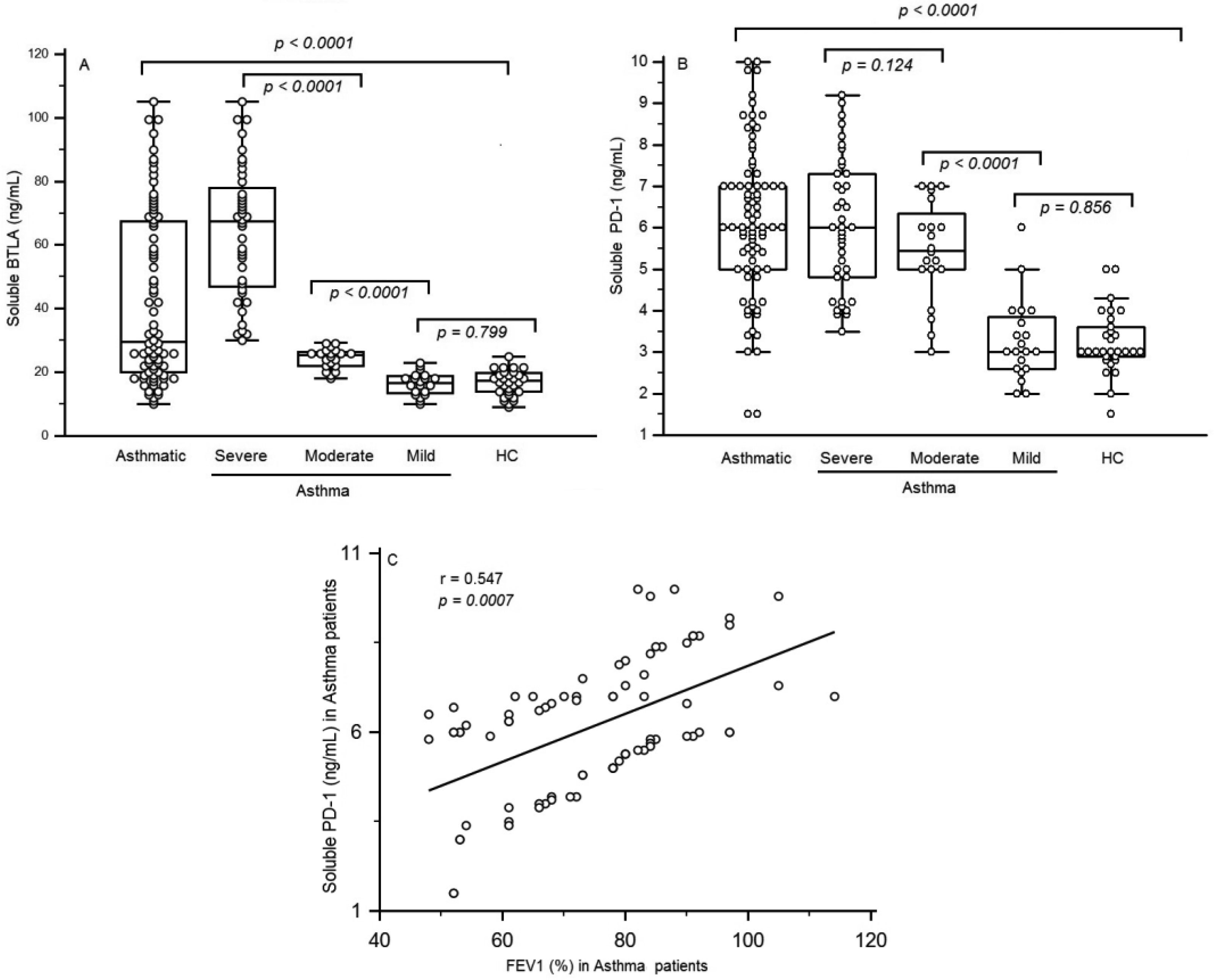
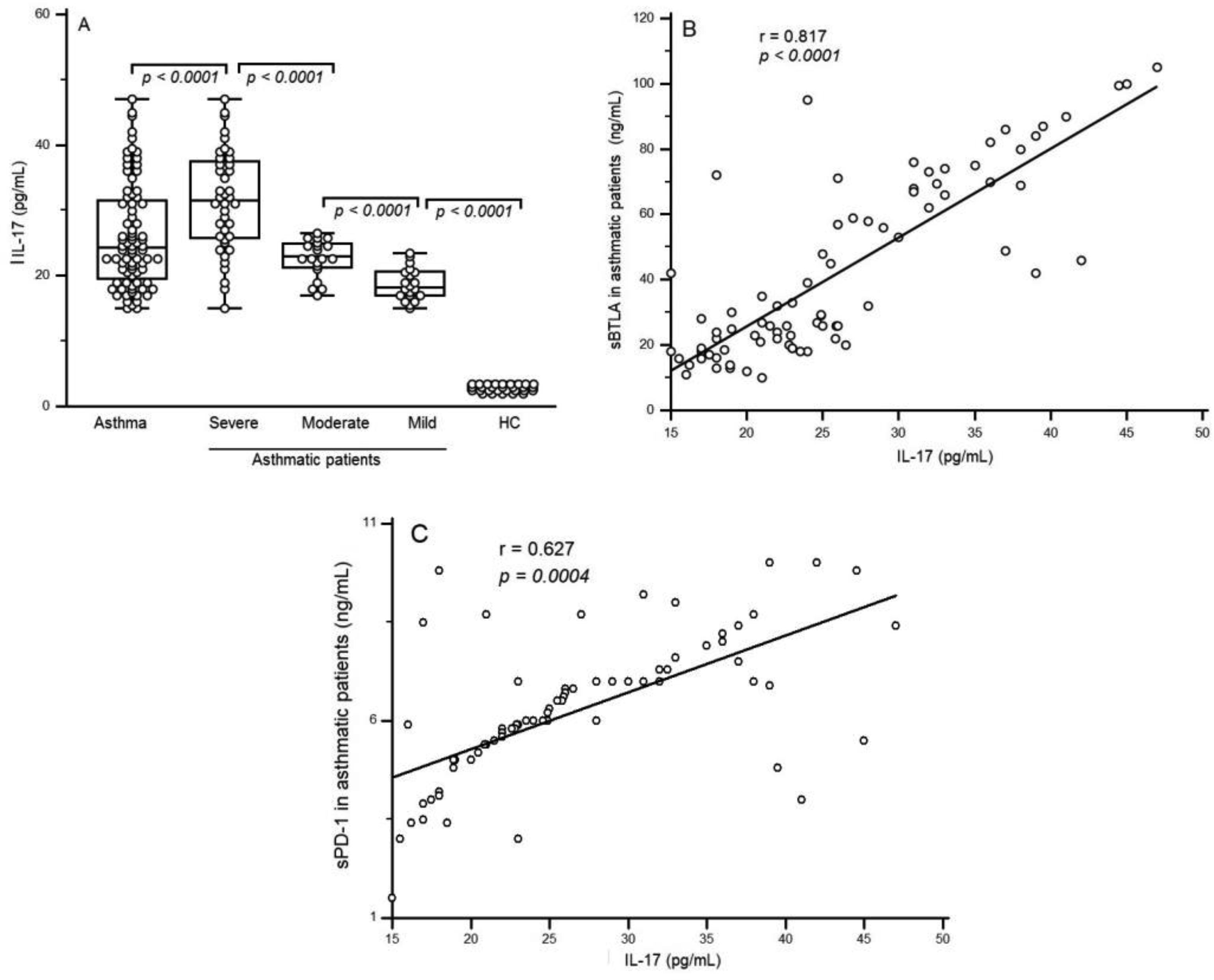
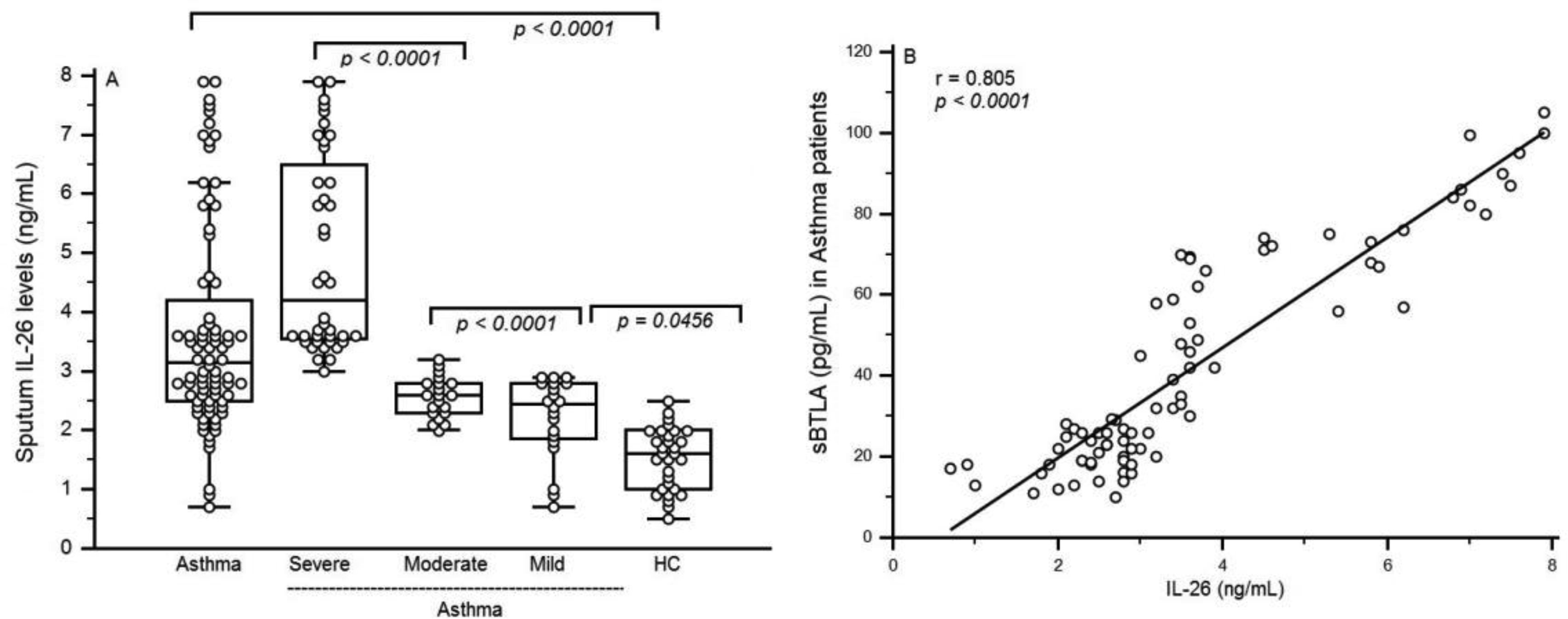
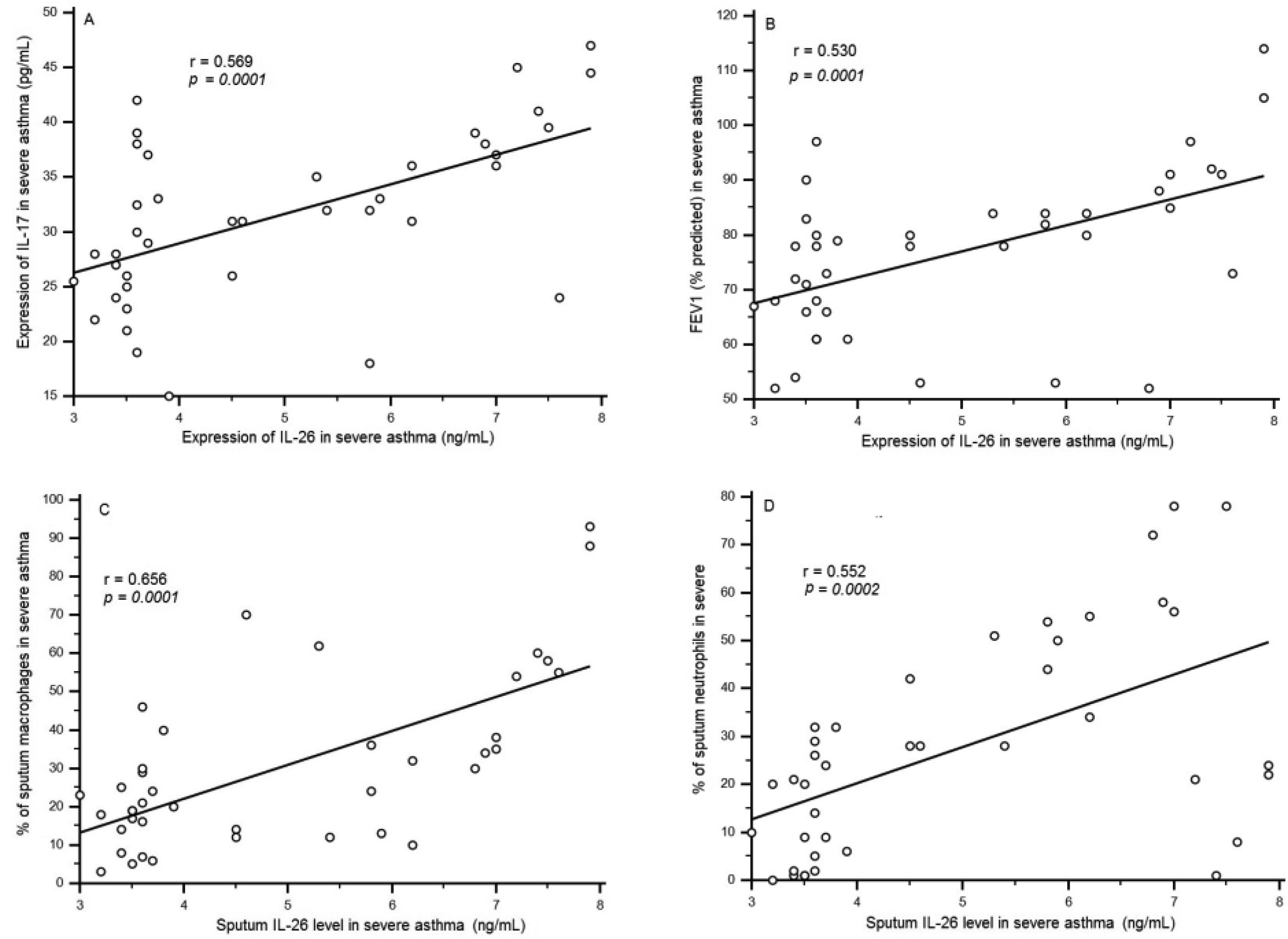
Figures(4) / Tables(2)

Sabrine Louhaichi, Mariem Salhi, Anissa Berraïes, Besma Hamdi, Jamel Ammar, Kamel Hamzaoui, Agnès Hamzaoui. Co-inhibitory receptors in female asthmatic patients: Correlation with IL-17 and IL-26[J]. AIMS Allergy and Immunology, 2018, 2(1): 10-23. doi: 10.3934/Allergy.2018.1.10







 DownLoad:
DownLoad: